

Autocrine IFN-γ promotes naive CD8 T cell differentiation and synergizes with IFN-α to stimulate strong function
- PMID: 22706089
- PMCID: PMC3392455
- DOI: 10.4049/jimmunol.1102727
Autocrine IFN-γ promotes naive CD8 T cell differentiation and synergizes with IFN-α to stimulate strong function
Abstract
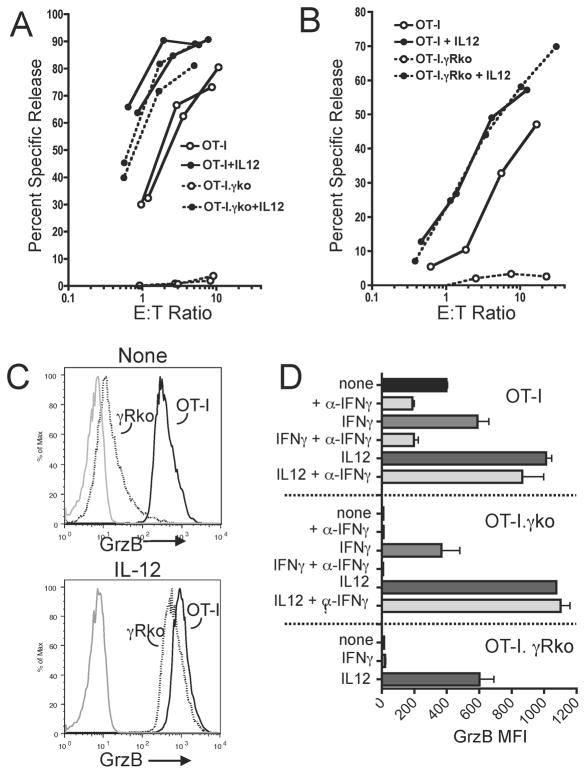
Autocrine IFN-γ signaling is important for CD4 differentiation to Th1 effector cells, but it has been unclear whether it contributes to CD8 T cell differentiation. We show in this paper that naive murine CD8 T cells rapidly and transiently produce low levels of IFN-γ upon stimulation with Ag and B7-1, with production peaking at ∼8 h and declining by 24 h. The autocrine IFN-γ signals for upregulation of expression of T-bet and granzyme B and induces weak cytolytic activity and effector IFN-γ production. IFN-α acts synergistically with IFN-γ to support development of strong effector functions, whereas IL-12 induces high T-bet expression and strong function in the absence of IFN-γ signaling. Thus, IFN-γ is not only an important CD8 T cell effector cytokine, it is an autocrine/paracrine factor whose contributions to differentiation vary depending on whether the response is supported by IL-12 or type I IFN.
Figures








References
-
- Aichele P, Unsoeld H, Koshella M, Schweier O, Kalinke U, Vucikuja S. Cutting Edge: CD8 T cells specific for lymphocytic choriomeningitis virus require Type I IFN receptor for clonal expansion. J Immunol. 2006;176:4525–4529. - PubMed
-
- Curtsinger J, Gerner MY, Lins DC, Mescher MF. Signal 3 availability limits the CD8 T cell response to a solid tumor. J Immunol. 2007;178:6752–6760. - PubMed
-
- Filatenkov AA, Jacovetty EL, Fischer UB, Curtsinger JM, Mescher MF, Ingulli E. CD4 T Cell-Dependent Conditioning of Dendritic Cells to Produce IL-12 Results in CD8-Mediated Graft Rejection and Avoidance of Tolerance. J Immunol. 2005;174:6909–6917. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
