

Physical confinement alters tumor cell adhesion and migration phenotypes
- PMID: 22707566
- PMCID: PMC3448771
- DOI: 10.1096/fj.12-211441
Physical confinement alters tumor cell adhesion and migration phenotypes
Abstract
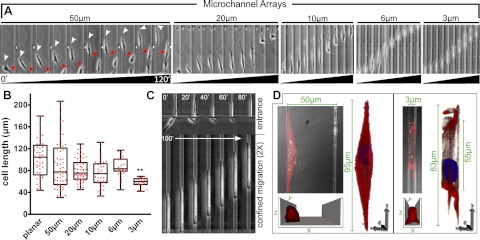
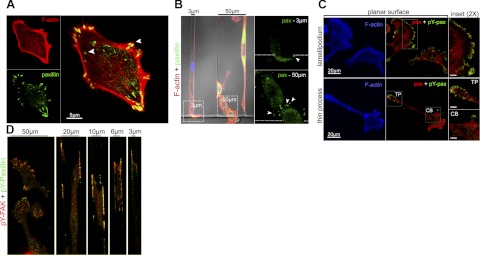
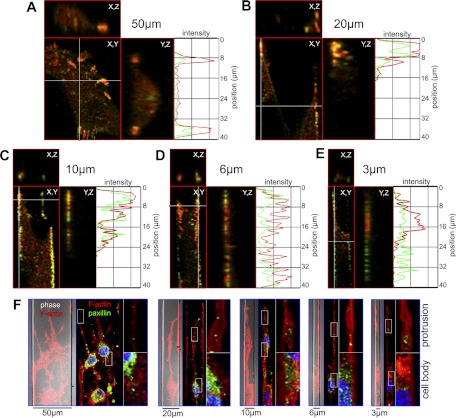
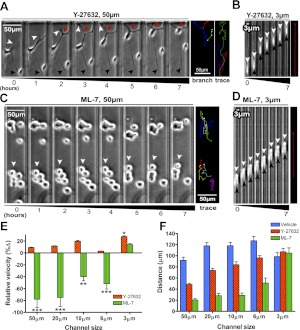
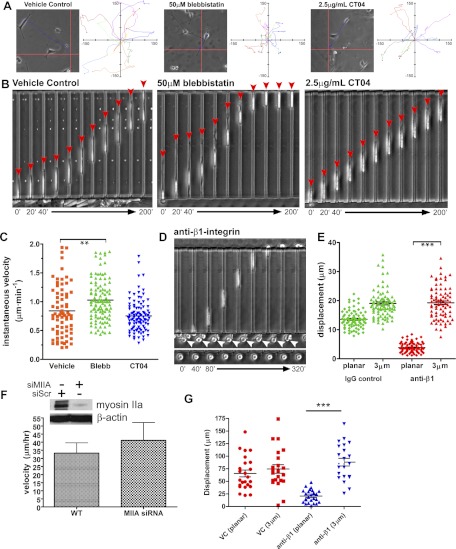
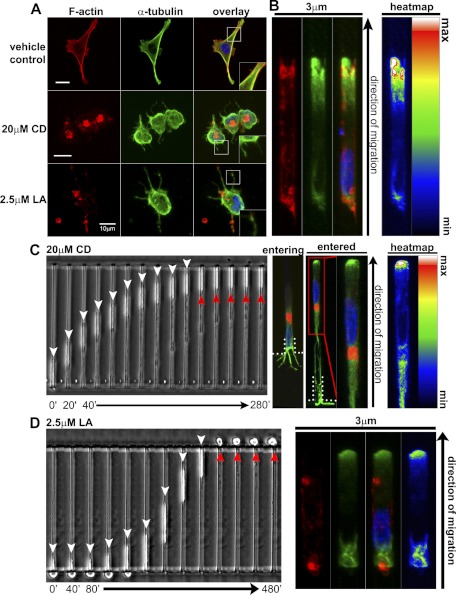
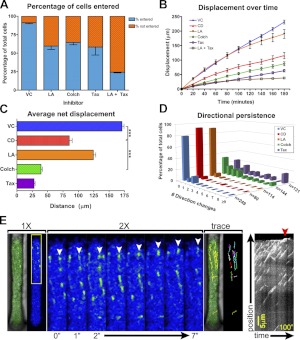
Cell migration on planar surfaces is driven by cycles of actin protrusion, integrin-mediated adhesion, and myosin-mediated contraction; however, this mechanism may not accurately describe movement in 3-dimensional (3D) space. By subjecting cells to restrictive 3D environments, we demonstrate that physical confinement constitutes a biophysical stimulus that alters cell morphology and suppresses mesenchymal motility in human breast carcinoma (MDA-MB-231). Dorsoventral polarity, stress fibers, and focal adhesions are markedly attenuated by confinement. Inhibitors of myosin, Rho/ROCK, or β1-integrins do not impair migration through 3-μm-wide channels (confinement), even though these treatments repress motility in 50-μm-wide channels (unconfined migration) by ≥50%. Strikingly, confined migration persists even when F-actin is disrupted, but depends largely on microtubule (MT) dynamics. Interfering with MT polymerization/depolymerization causes confined cells to undergo frequent directional changes, thereby reducing the average net displacement by ≥80% relative to vehicle controls. Live-cell EB1-GFP imaging reveals that confinement redirects MT polymerization toward the leading edge, where MTs continuously impact during advancement of the cell front. These results demonstrate that physical confinement can induce cytoskeletal alterations that reduce the dependence of migrating cells on adhesion-contraction force coupling. This mechanism may explain why integrins can exhibit reduced or altered function during migration in 3D environments.
Figures
References
-
- Even-Ram S., Yamada K. M. (2005) Cell migration in 3D matrix. Curr. Opin. Cell Biol. 17, 524–532 - PubMed
-
- Lakshman N., Kim A., Bayless K. J., Davis G. E., Petroll W. M. (2007) Rho plays a central role in regulating local cell-matrix mechanical interactions in 3D culture. Cell Motil. Cytoskeleton 64, 434–445 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
