

The folliculin-FNIP1 pathway deleted in human Birt-Hogg-Dubé syndrome is required for murine B-cell development
- PMID: 22709692
- PMCID: PMC3418720
- DOI: 10.1182/blood-2012-02-410407
The folliculin-FNIP1 pathway deleted in human Birt-Hogg-Dubé syndrome is required for murine B-cell development
Abstract
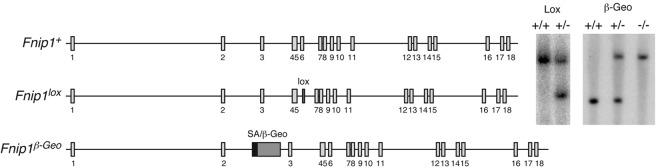
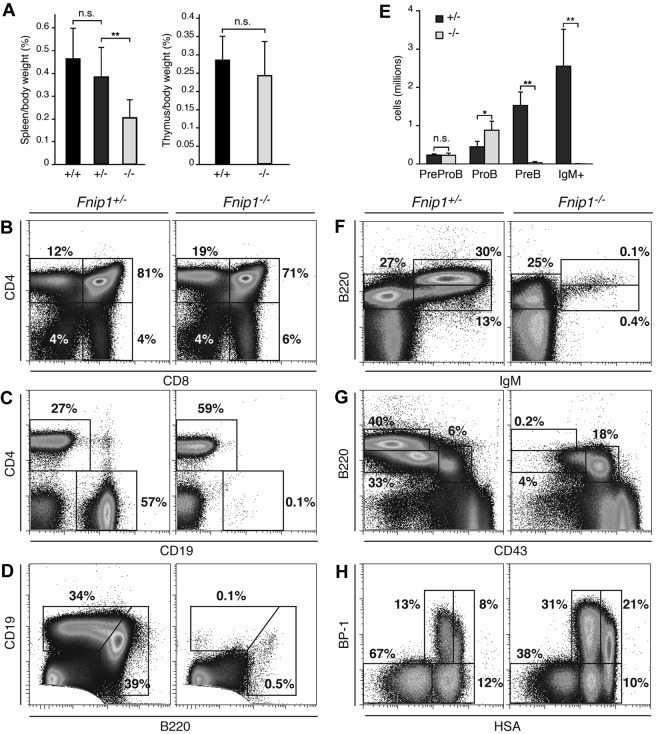
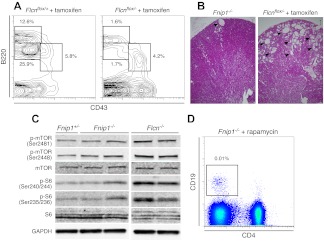
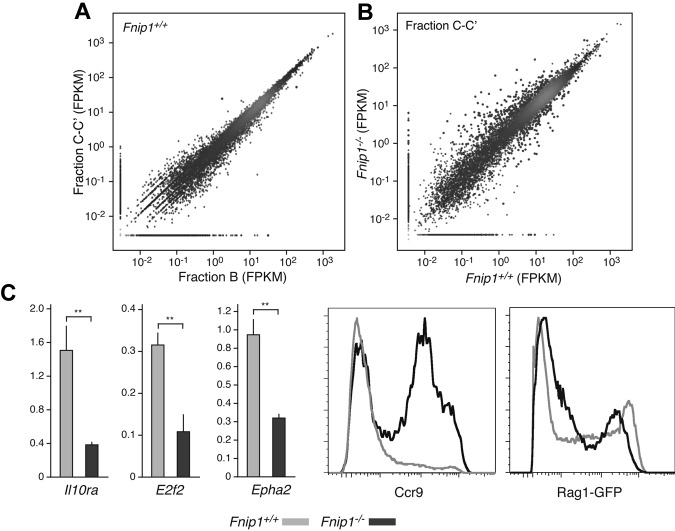
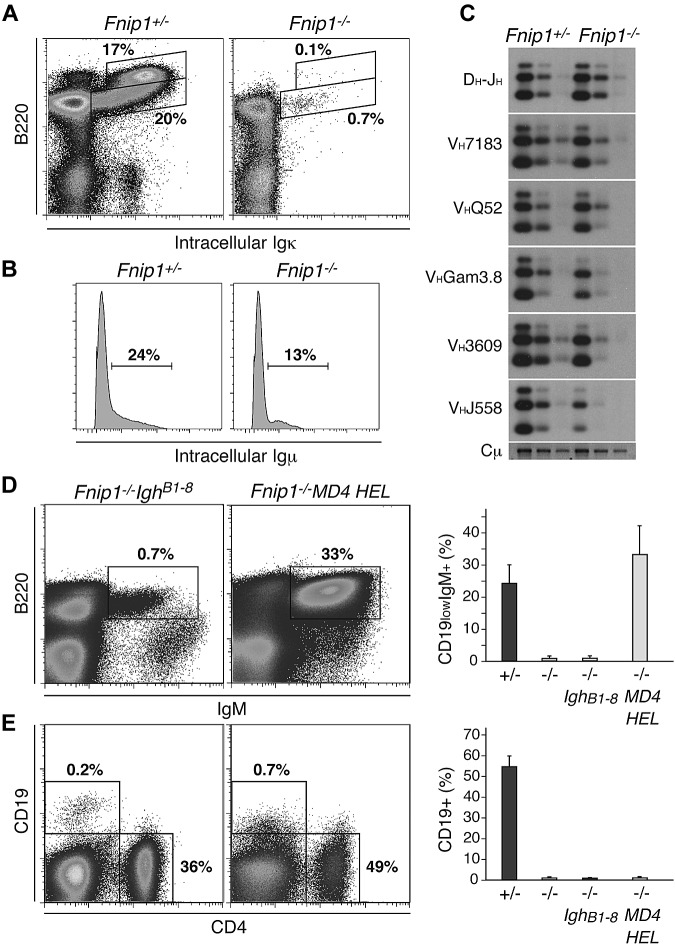
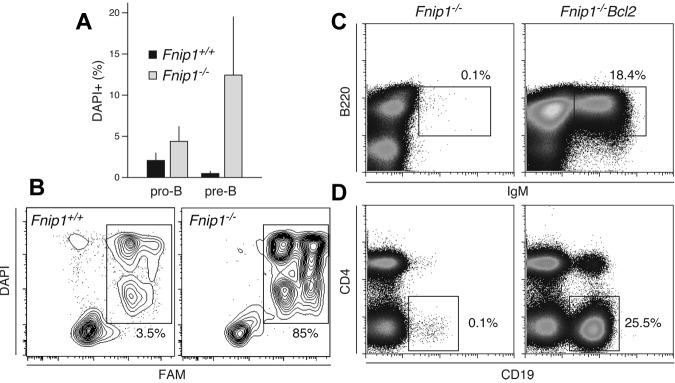
Birt-Hogg-Dubé (BHD) syndrome is an autosomal dominant disorder characterized by cutaneous fibrofolliculomas, pulmonary cysts, and kidney malignancies. Affected individuals carry germ line mutations in folliculin (FLCN), a tumor suppressor gene that becomes biallelically inactivated in kidney tumors by second-hit mutations. Similar to other factors implicated in kidney cancer, FLCN has been shown to modulate activation of mammalian target of rapamycin (mTOR). However, its precise in vivo function is largely unknown because germ line deletion of Flcn results in early embryonic lethality in animal models. Here, we describe mice deficient in the newly characterized folliculin-interacting protein 1 (Fnip1). In contrast to Flcn, Fnip1(-/-) mice develop normally, are not susceptible to kidney neoplasia, but display a striking pro-B cell block that is entirely independent of mTOR activity. We show that this developmental arrest results from rapid caspase-induced pre-B cell death, and that a Bcl2 transgene reconstitutes mature B-cell populations, respectively. We also demonstrate that conditional deletion of Flcn recapitulates the pro-B cell arrest of Fnip1(-/-) mice. Our studies thus demonstrate that the FLCN-FNIP complex deregulated in BHD syndrome is absolutely required for B-cell differentiation, and that it functions through both mTOR-dependent and independent pathways.
Figures
References
-
- Nickerson ML, Warren MB, Toro JR, et al. Mutations in a novel gene lead to kidney tumors, lung wall defects, and benign tumors of the hair follicle in patients with the Birt-Hogg-Dube syndrome. Cancer Cell. 2002;2(2):157–164. - PubMed
-
- Okimoto K, Kouchi M, Matsumoto I, Sakurai J, Kobayashi T, Hino O. Natural history of the Nihon rat model of BHD. Curr Mol Med. 2004;4(8):887–893. - PubMed
-
- Vocke CD, Yang Y, Pavlovich CP, et al. High frequency of somatic frameshift BHD gene mutations in Birt-Hogg-Dube-associated renal tumors. J Natl Cancer Inst. 2005;97(12):931–935. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
