

A novel membrane-dependent on/off switch mechanism of talin FERM domain at sites of cell adhesion
- PMID: 22710802
- PMCID: PMC3494399
- DOI: 10.1038/cr.2012.97
A novel membrane-dependent on/off switch mechanism of talin FERM domain at sites of cell adhesion
Abstract
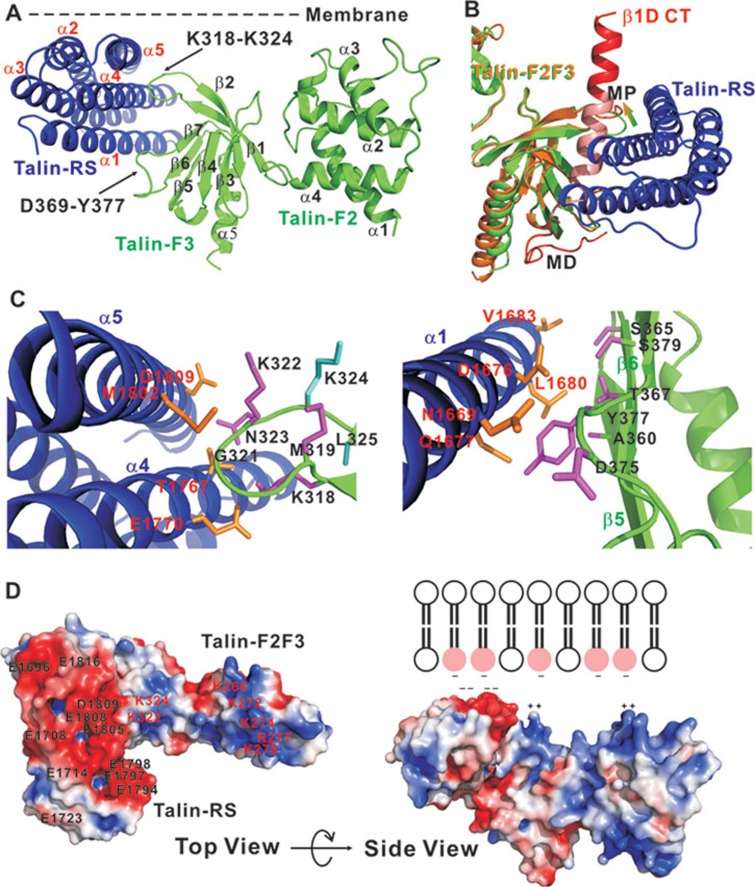
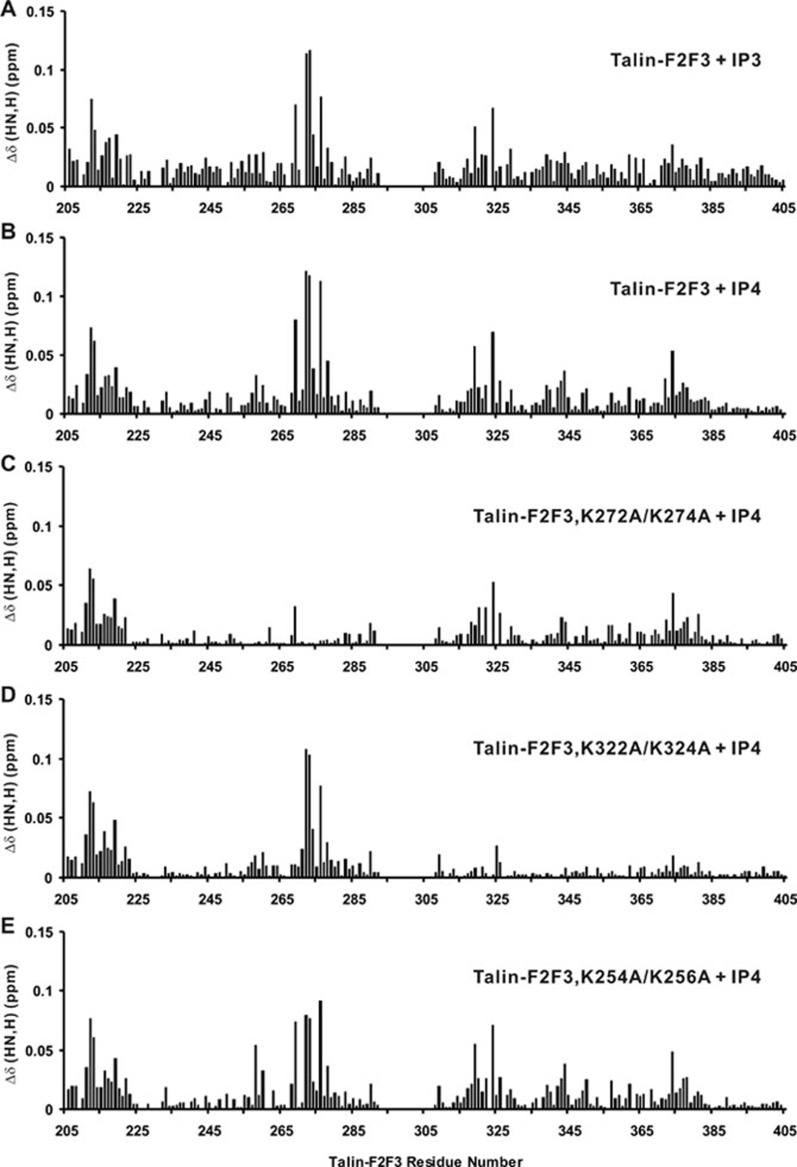
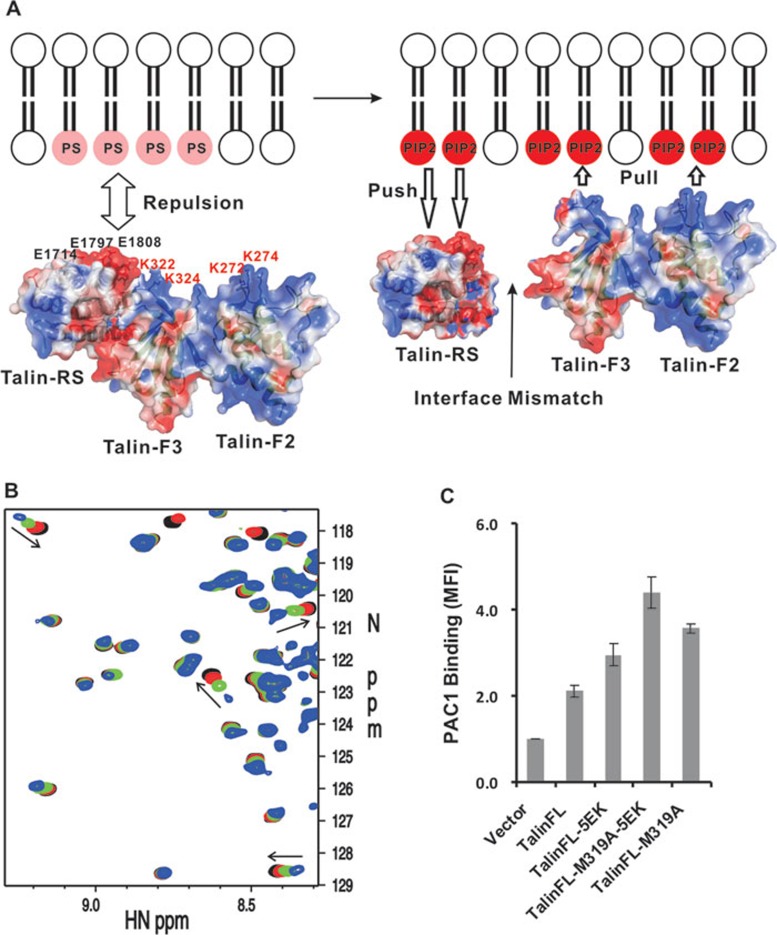
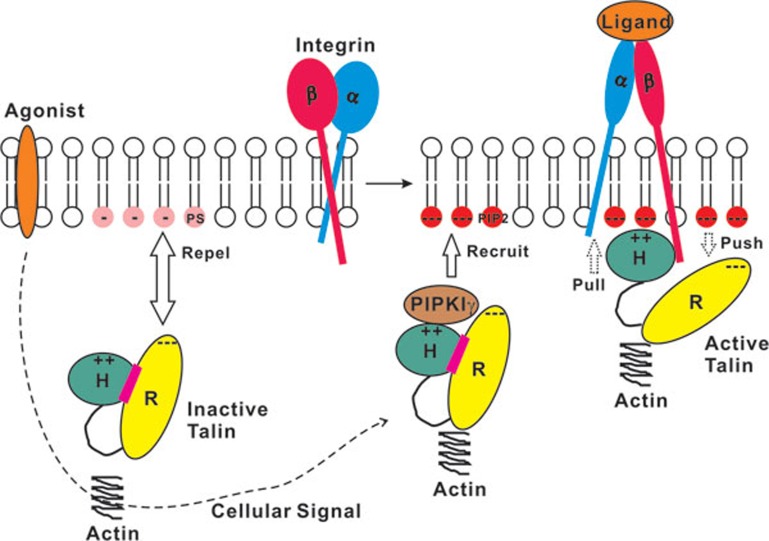
The activation of heterodimeric (α/β) integrin transmembrane receptors by cytosolic protein talin is crucial for regulating diverse cell-adhesion-dependent processes, including blood coagulation, tissue remodeling, and cancer metastasis. This process is triggered by the coincident binding of N-terminal FERM (four-point-one-protein/ezrin/radixin/moesin) domain of talin (talin-FERM) to the inner membrane surface and integrin β cytoplasmic tail, but how these binding events are spatiotemporally regulated remains obscure. Here we report the crystal structure of a dormant talin, revealing how a C-terminal talin rod segment (talin-RS) self-masks a key integrin-binding site on talin-FERM via a large interface. Unexpectedly, the structure also reveals a distinct negatively charged surface on talin-RS that electrostatically hinders the talin-FERM binding to the membrane. Such a dual inhibitory topology for talin is consistent with the biochemical and functional data, but differs significantly from a previous model. We show that upon enrichment with phosphotidylinositol-4,5-bisphosphate (PIP2) - a known talin activator, membrane strongly attracts a positively charged surface on talin-FERM and simultaneously repels the negatively charged surface on talin-RS. Such an electrostatic "pull-push" process promotes the relief of the dual inhibition of talin-FERM, which differs from the classic "steric clash" model for conventional PIP2-induced FERM domain activation. These data therefore unravel a new type of membrane-dependent FERM domain regulation and illustrate how it mediates the talin on/off switches to regulate integrin transmembrane signaling and cell adhesion.
Figures
Similar articles
-
Pull and push: talin activation for integrin signaling.Cell Res. 2012 Nov;22(11):1512-4. doi: 10.1038/cr.2012.103. Epub 2012 Jul 10. Cell Res. 2012. PMID: 22777425 Free PMC article.
-
A conserved lipid-binding loop in the kindlin FERM F1 domain is required for kindlin-mediated αIIbβ3 integrin coactivation.J Biol Chem. 2012 Mar 2;287(10):6979-90. doi: 10.1074/jbc.M111.330845. Epub 2012 Jan 10. J Biol Chem. 2012. PMID: 22235127 Free PMC article.
-
Affinity of talin-1 for the β3-integrin cytosolic domain is modulated by its phospholipid bilayer environment.Proc Natl Acad Sci U S A. 2012 Jan 17;109(3):793-8. doi: 10.1073/pnas.1117220108. Epub 2011 Dec 30. Proc Natl Acad Sci U S A. 2012. PMID: 22210111 Free PMC article.
-
The talin-tail interaction places integrin activation on FERM ground.Trends Biochem Sci. 2004 Aug;29(8):429-35. doi: 10.1016/j.tibs.2004.06.005. Trends Biochem Sci. 2004. PMID: 15362227 Review.
-
Talin controls integrin activation.Biochem Soc Trans. 2004 Jun;32(Pt3):434-7. doi: 10.1042/BST0320434. Biochem Soc Trans. 2004. PMID: 15157154 Review.
Cited by
-
β2 Integrin Signaling Cascade in Neutrophils: More Than a Single Function.Front Immunol. 2021 Feb 18;11:619925. doi: 10.3389/fimmu.2020.619925. eCollection 2020. Front Immunol. 2021. PMID: 33679708 Free PMC article. Review.
-
Erythrocyte membrane protein band 4.1-like 3 inhibits osteosarcoma cell invasion through regulation of Snai1-induced epithelial-to-mesenchymal transition.Aging (Albany NY). 2020 Dec 11;13(2):1947-1961. doi: 10.18632/aging.202158. Epub 2020 Dec 11. Aging (Albany NY). 2020. PMID: 33323539 Free PMC article.
-
Adhesions Assemble!-Autoinhibition as a Major Regulatory Mechanism of Integrin-Mediated Adhesion.Front Mol Biosci. 2019 Dec 17;6:144. doi: 10.3389/fmolb.2019.00144. eCollection 2019. Front Mol Biosci. 2019. PMID: 31921890 Free PMC article. Review.
-
Selectins and chemokines use shared and distinct signals to activate β2 integrins in neutrophils.Blood Adv. 2018 Apr 10;2(7):731-744. doi: 10.1182/bloodadvances.2017015602. Blood Adv. 2018. PMID: 29592875 Free PMC article.
-
Talin autoinhibition is required for morphogenesis.Curr Biol. 2013 Sep 23;23(18):1825-33. doi: 10.1016/j.cub.2013.07.054. Epub 2013 Sep 5. Curr Biol. 2013. PMID: 24012314 Free PMC article.
References
-
- Critchley DR. Biochemical and structural properties of the integrin-associated cytoskeletal protein talin. Annu Rev Biophys. 2009;38:235–254. - PubMed
-
- Moser M, Legate KR, Zent R, Fassler R. The tail of integrins, talin, and kindlins. Science. 2009;324:895–899. - PubMed
-
- Wehrle-Haller B. Structure and function of focal adhesions. Curr Opin Cell Biol. 2012;24:116–124. - PubMed
-
- Turner CE, Burridge K. Transmembrane molecular assemblies in cell-extracellular matrix interactions. Curr Opin Cell Biol. 1991;3:849–853. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
