

γ-Aminobutyric acid type A (GABAA) receptor α subunits play a direct role in synaptic versus extrasynaptic targeting
- PMID: 22711532
- PMCID: PMC3431651
- DOI: 10.1074/jbc.M112.360461
γ-Aminobutyric acid type A (GABAA) receptor α subunits play a direct role in synaptic versus extrasynaptic targeting
Abstract
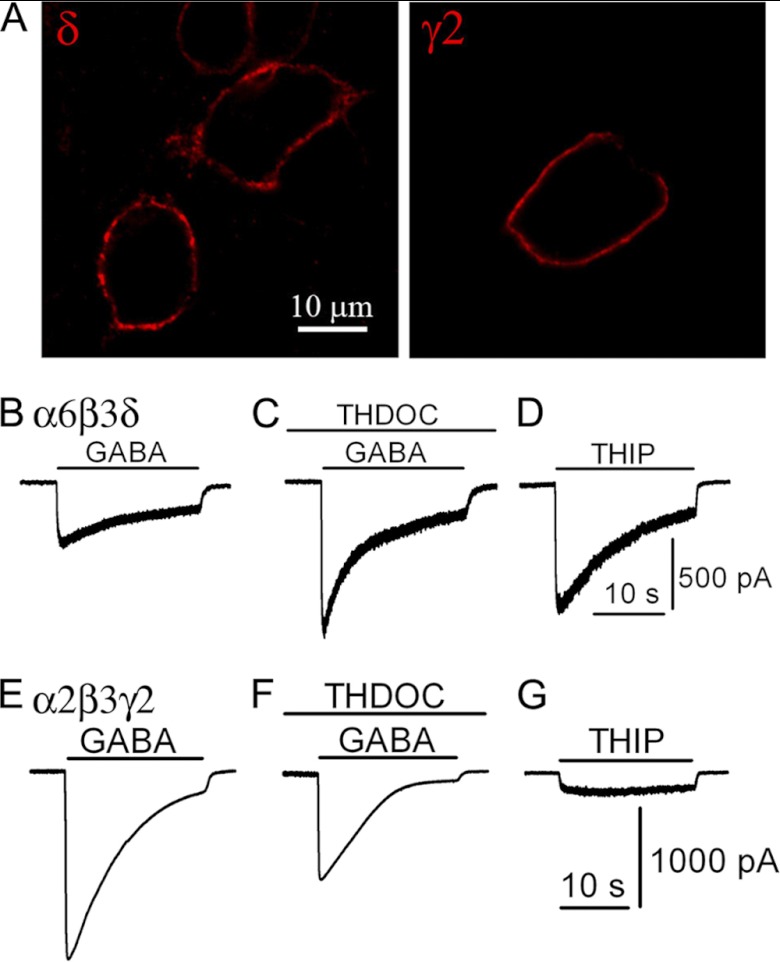
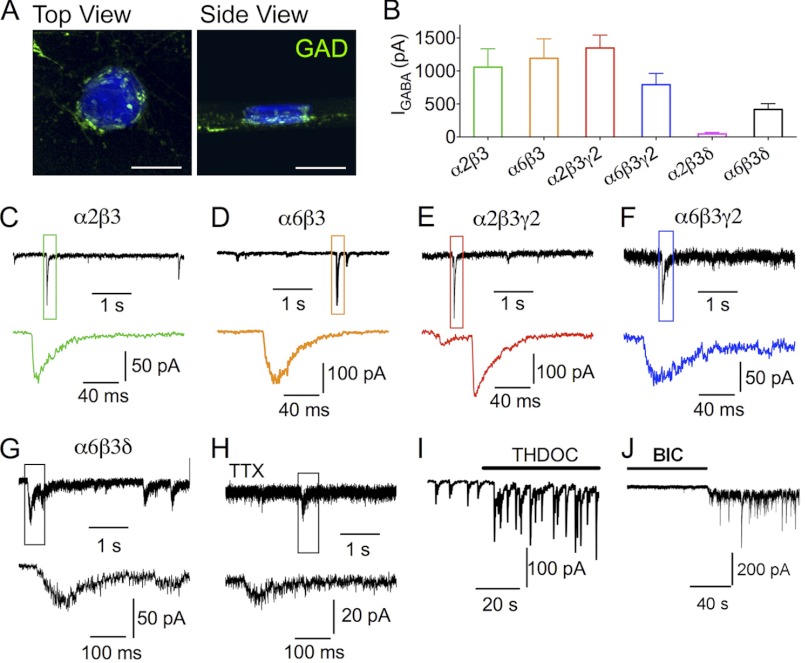
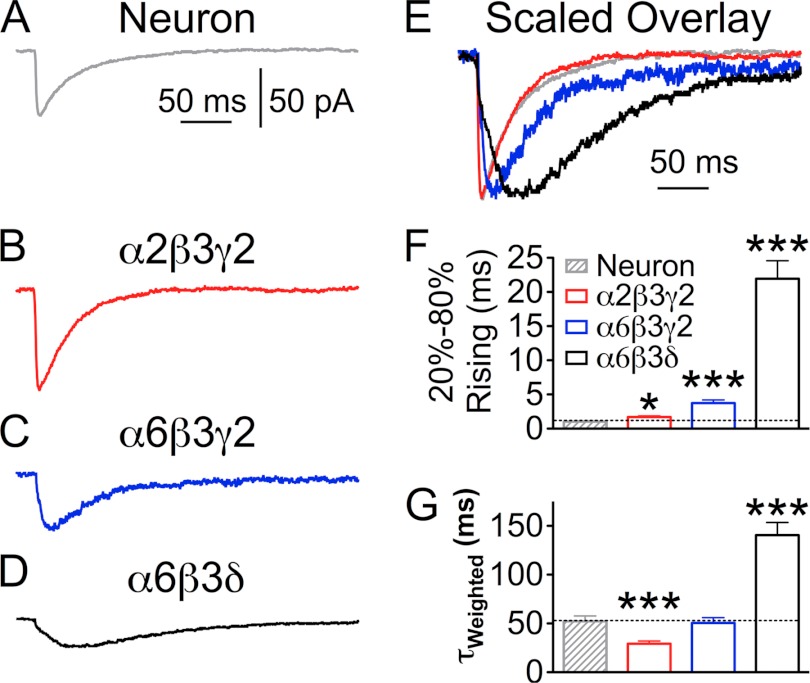
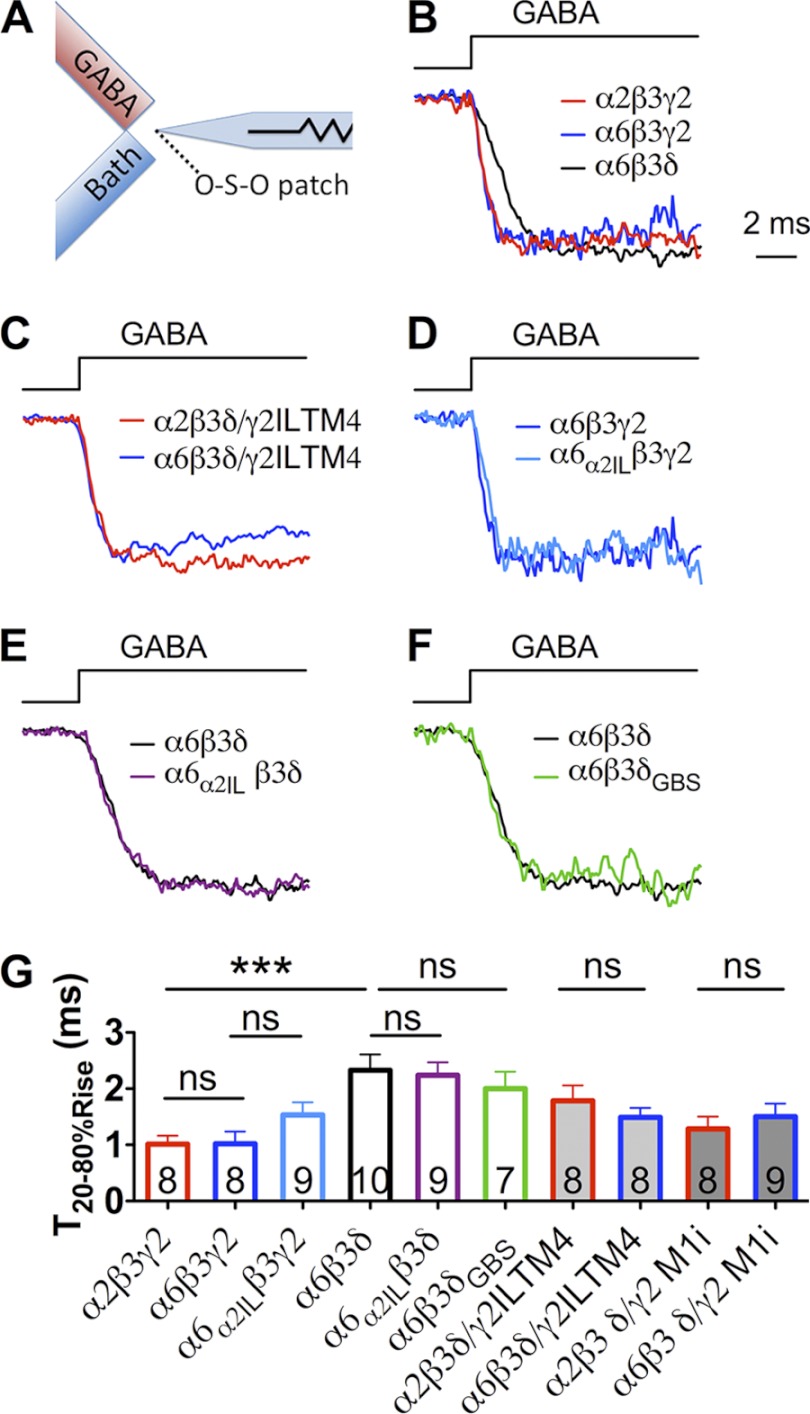
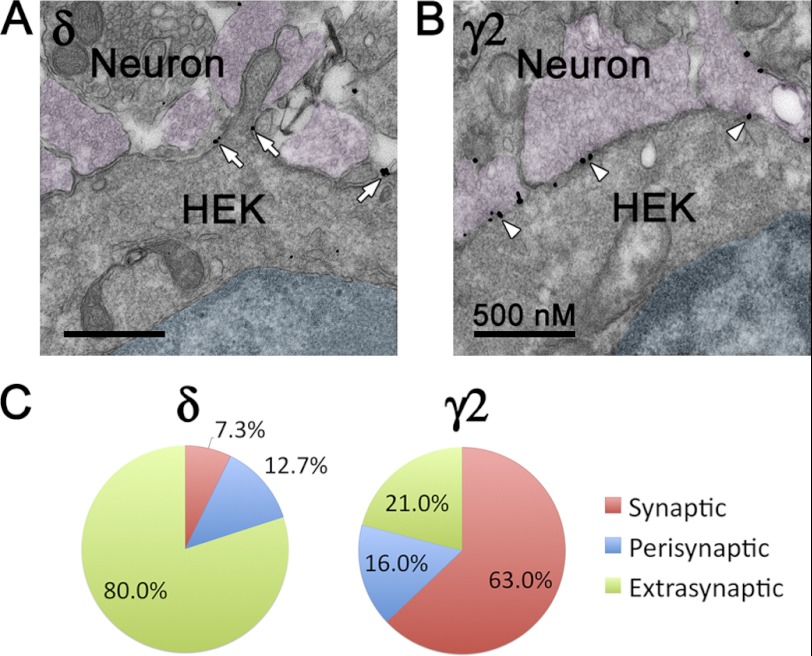
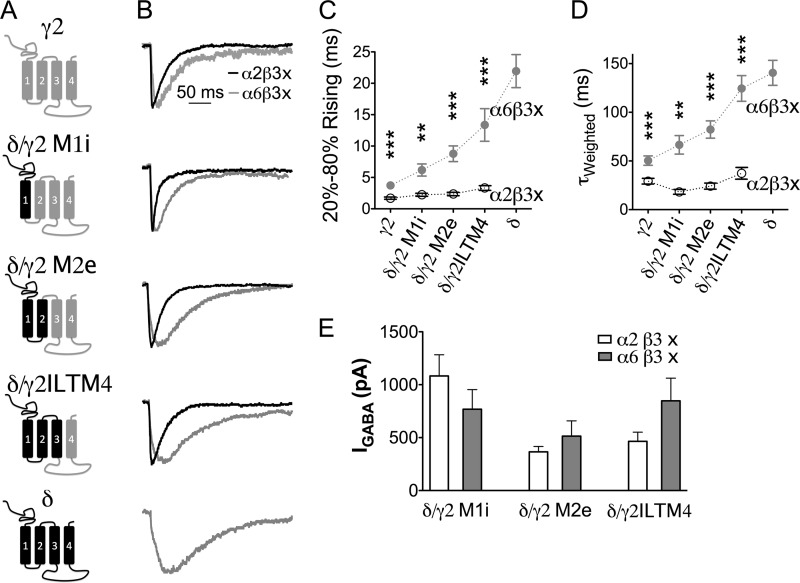
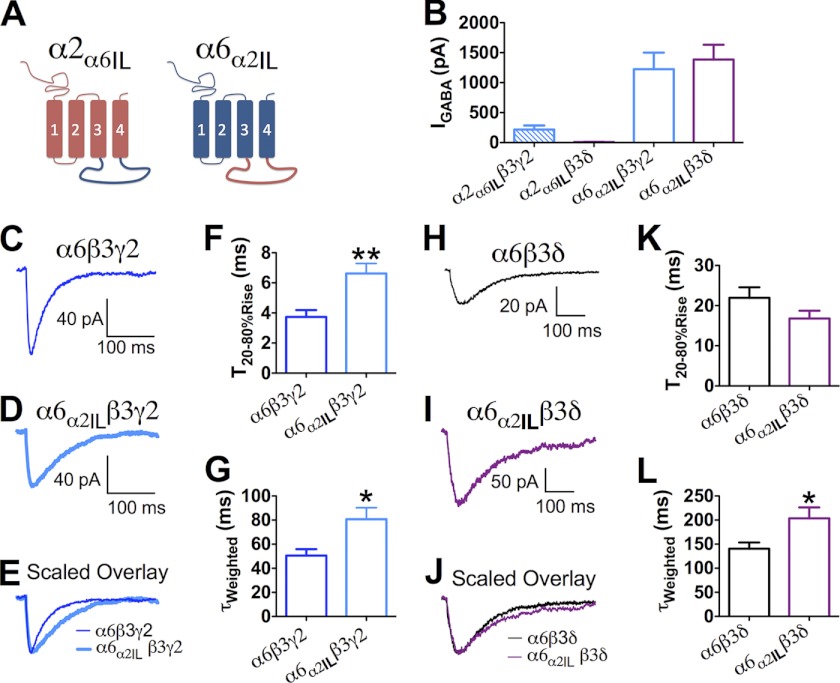
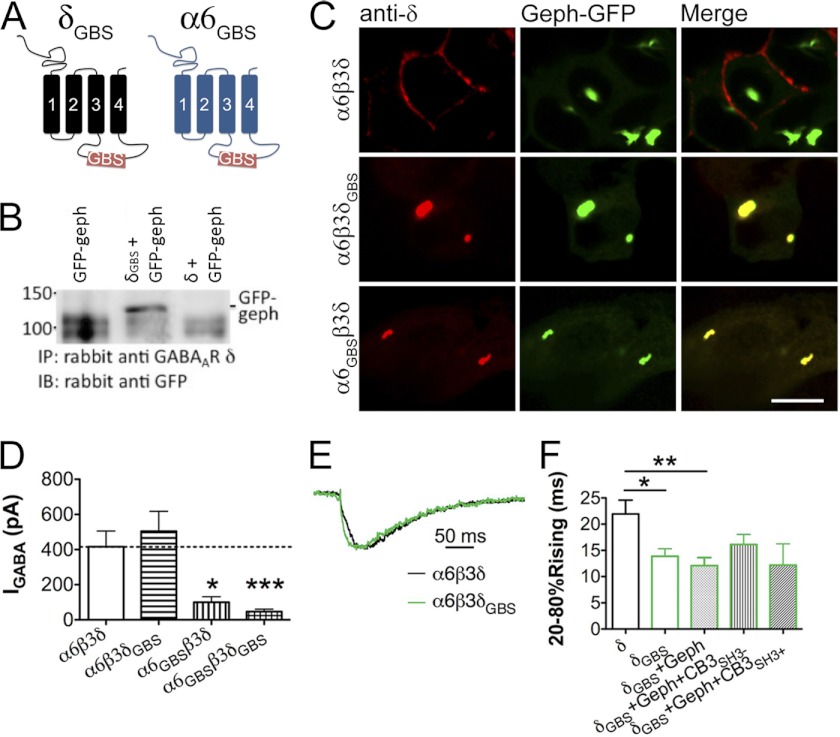
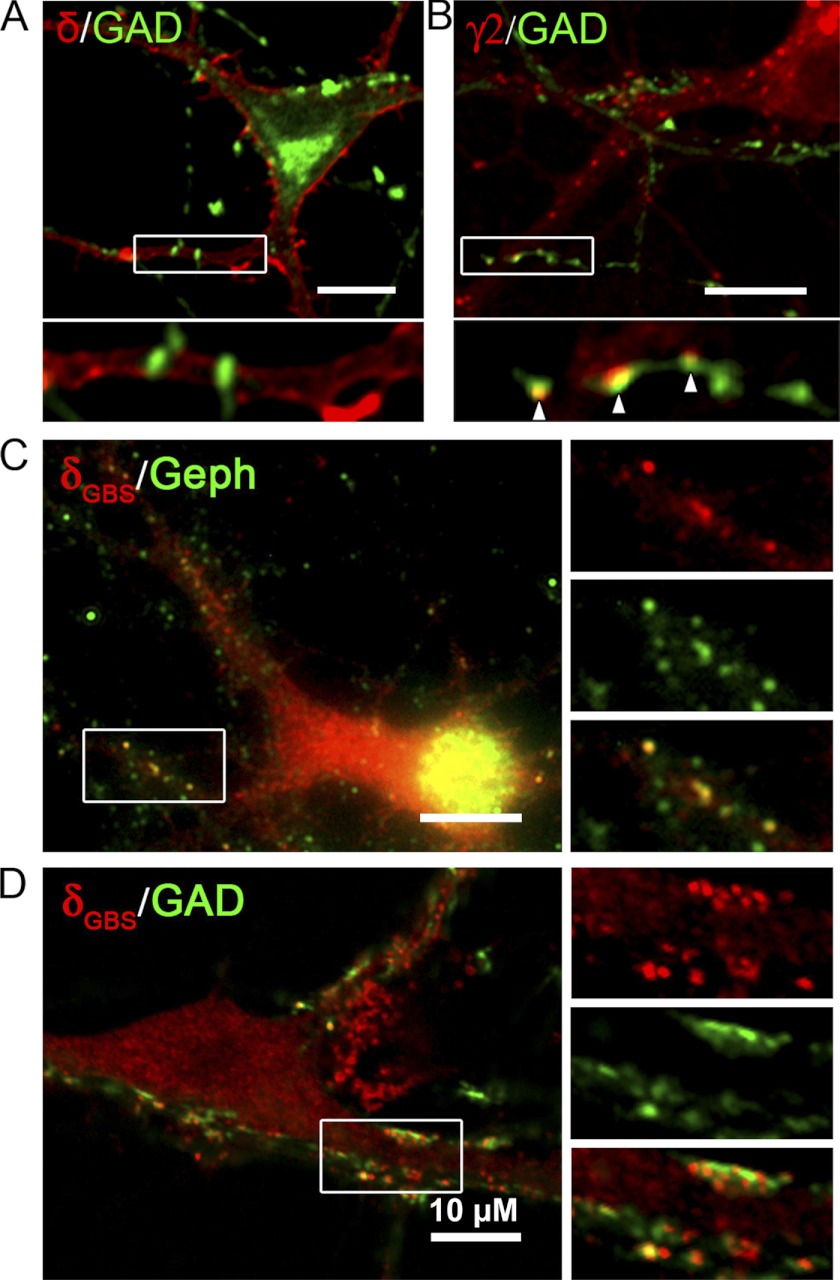
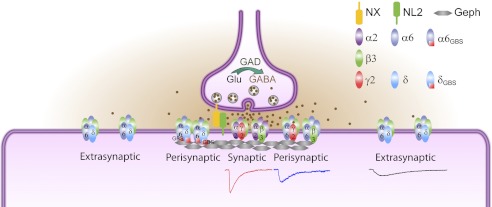
GABA(A) receptors (GABA(A)-Rs) are localized at both synaptic and extrasynaptic sites, mediating phasic and tonic inhibition, respectively. Previous studies suggest an important role of γ2 and δ subunits in synaptic versus extrasynaptic targeting of GABA(A)-Rs. Here, we demonstrate differential function of α2 and α6 subunits in guiding the localization of GABA(A)-Rs. To study the targeting of specific subtypes of GABA(A)-Rs, we used a molecularly engineered GABAergic synapse model to precisely control the GABA(A)-R subunit composition. We found that in neuron-HEK cell heterosynapses, GABAergic events mediated by α2β3γ2 receptors were very fast (rise time ∼2 ms), whereas events mediated by α6β3δ receptors were very slow (rise time ∼20 ms). Such an order of magnitude difference in rise time could not be attributed to the minute differences in receptor kinetics. Interestingly, synaptic events mediated by α6β3 or α6β3γ2 receptors were significantly slower than those mediated by α2β3 or α2β3γ2 receptors, suggesting a differential role of α subunit in receptor targeting. This was confirmed by differential targeting of the same δ-γ2 chimeric subunits to synaptic or extrasynaptic sites, depending on whether it was co-assembled with the α2 or α6 subunit. In addition, insertion of a gephyrin-binding site into the intracellular domain of α6 and δ subunits brought α6β3δ receptors closer to synaptic sites. Therefore, the α subunits, together with the γ2 and δ subunits, play a critical role in governing synaptic versus extrasynaptic targeting of GABA(A)-Rs, possibly through differential interactions with gephyrin.
Figures
References
-
- Sigel E., Baur R., Boulineau N., Minier F. (2006) Impact of subunit positioning on GABAA receptor function. Biochem. Soc. Trans. 34, 868–871 - PubMed
-
- Farrant M., Nusser Z. (2005) Variations on an inhibitory theme. Phasic and tonic activation of GABAA receptors. Nat. Rev. Neurosci. 6, 215–229 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- RC1 MH089111/MH/NIMH NIH HHS/United States
- R01 NS051590/NS/NINDS NIH HHS/United States
- NS38752/NS/NINDS NIH HHS/United States
- R01 MH062391/MH/NIMH NIH HHS/United States
- R01 NS033300/NS/NINDS NIH HHS/United States
- R56 MH062391/MH/NIMH NIH HHS/United States
- R21 NS054858/NS/NINDS NIH HHS/United States
- R01 MH083911/MH/NIMH NIH HHS/United States
- R01 NS038752/NS/NINDS NIH HHS/United States
- MH062391/MH/NIMH NIH HHS/United States
- NS054858/NS/NINDS NIH HHS/United States
- MH083911/MH/NIMH NIH HHS/United States
- NS33300/NS/NINDS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
