

Cold-inducible RNA-binding protein (Cirp) interacts with Dyrk1b/Mirk and promotes proliferation of immature male germ cells in mice
- PMID: 22711815
- PMCID: PMC3390833
- DOI: 10.1073/pnas.1121524109
Cold-inducible RNA-binding protein (Cirp) interacts with Dyrk1b/Mirk and promotes proliferation of immature male germ cells in mice
Abstract
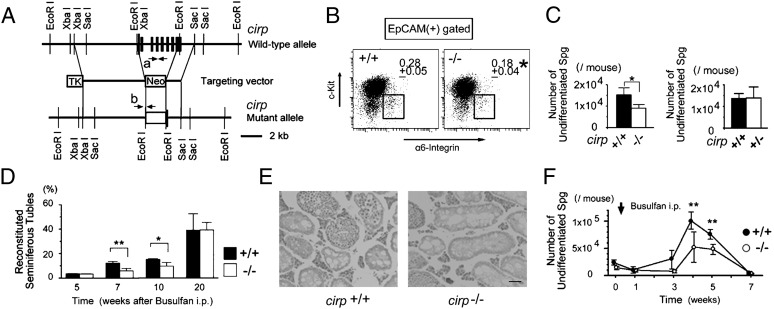
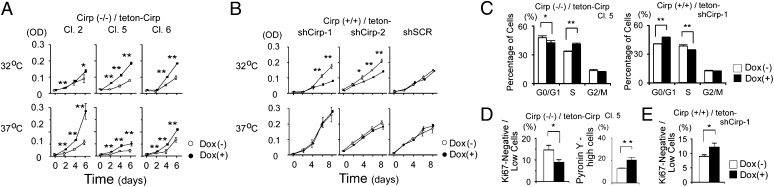
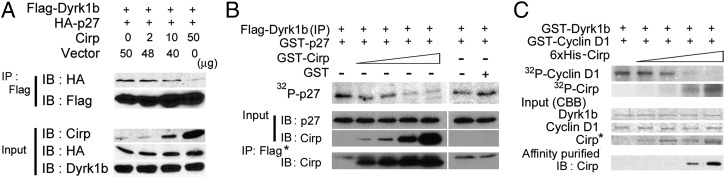
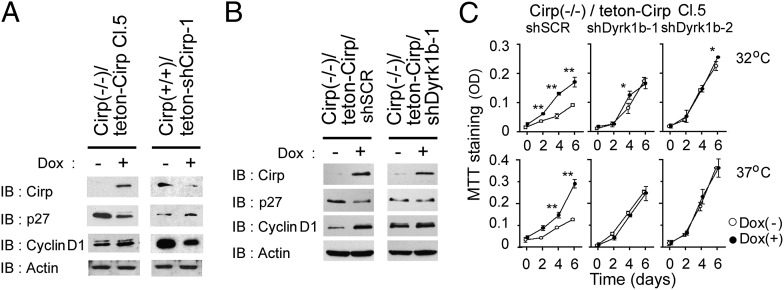
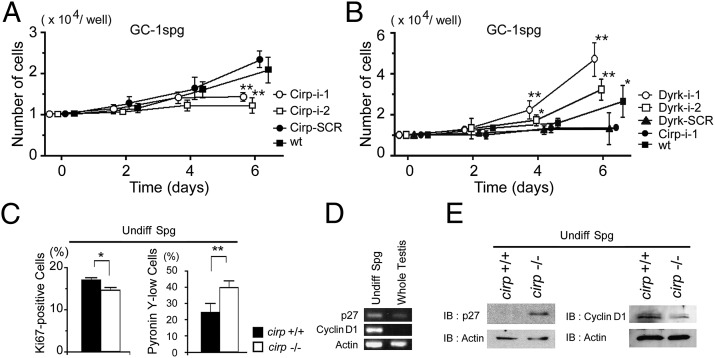
Cold-inducible RNA-binding protein (Cirp) was the first cold-shock protein identified in mammals. It is structurally quite different from bacterial cold-shock proteins and is induced in response to mild, but not severe, hypothermia. To clarify the physiological function of Cirp in vivo, we produced cirp-knockout mice. They showed neither gross abnormality nor defect in fertility, but the number of undifferentiated spermatogonia was significantly reduced and the recovery of spermatogenesis was delayed after treatment with a cytotoxic agent, busulfan. Cirp accelerated cell-cycle progression from G0 to G1 as well as from G1 to S phase in cultured mouse embryonic fibroblasts. Cirp directly bound to dual-specificity tyrosine-phosphorylation-regulated kinase 1B (Dyrk1b, also called Mirk) and inhibited its binding to p27, resulting in decreased phosphorylation and destabilization of p27. Cirp did not affect binding of Dyrk1b to cyclin D1 but inhibited phosphorylation of cyclin D1 by Dyrk1b, resulting in cyclin D1 stabilization. In the spermatogonial cell line GC-1spg, suppression of Cirp expression increased the protein level of p27, decreased that of cyclin D1, and decreased the growth rate, which depended on Dyrk1b. Consistent changes in the protein levels of p27 and cyclin D1 as well as the percentage of cells in G0 phase were observed in undifferentiated spermatogonia of cirp-knockout mice. In undifferentiated spermatogonia of wild-type mice, Cirp and Dyrk1b colocalized in the nucleus. Thus, our study demonstrates that Cirp functions to fine-tune the proliferation of undifferentiated spermatogonia by interacting with Dyrk1b.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Harrison RG, Weiner JS. Abdomino-testicular temperature gradients. J Physiol. 1948;107:48–49.
-
- Carrick F, Setchell B. The evolution of the scrotum. In: Calaby J, Tyndale-Biscoe T, editors. Reproduction and Evolution. Canberra: Australian Academy of Science; 1977. pp. 165–170.
-
- Fujita J. Cold shock response in mammalian cells. J Mol Microbiol Biotechnol. 1999;1:243–255. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
