

Characterization of secretory sphingomyelinase activity, lipoprotein sphingolipid content and LDL aggregation in ldlr-/- mice fed on a high-fat diet
- PMID: 22712892
- PMCID: PMC3475451
- DOI: 10.1042/BSR20120036
Characterization of secretory sphingomyelinase activity, lipoprotein sphingolipid content and LDL aggregation in ldlr-/- mice fed on a high-fat diet
Abstract
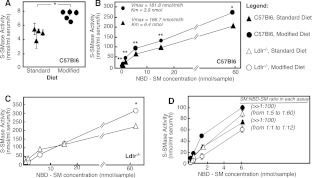
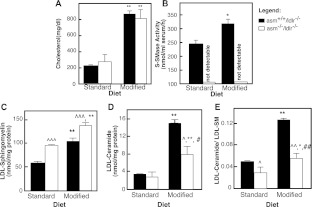
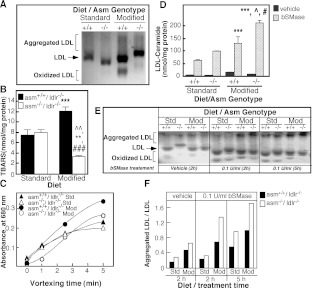
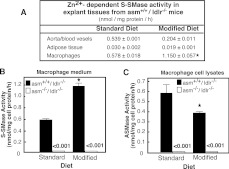
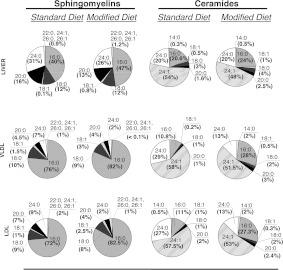
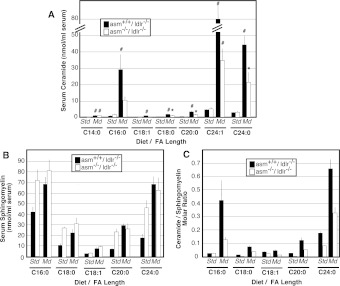
The propensity of LDLs (low-density lipoproteins) for aggregation and/or oxidation has been linked to their sphingolipid content, specifically the levels of SM (sphingomyelin) and ceramide. To investigate this association in vivo, ldlr (LDL receptor)-null mice (ldlr-/-) were fed on a modified (atherogenic) diet containing saturated fats and cholesterol. The diet led to significantly elevated SM content in all serum lipoproteins. In contrast, ceramide increased only in the LDL particles. MS-based analyses of the lipid acyl chain composition revealed a marked elevation in C16:0 fatty acid in SM and ceramide, consistent with the prevalence of palmitic acid in the modified diet. The diet also led to increased activity of the S-SMase [secretory SMase (sphingomyelinase)], a protein that is generated by ASMase (acid SMase) and acts on serum LDL. An increased macrophage secretion seemed to be responsible for the elevated S-SMase activity. ASMase-deficient mice (asm-/-/ldlr-/-) lacked S-SMase activity and were protected from diet-induced elevation in LDL ceramide. LDL from asm-/-/ldlr-/- mice fed on the modified diet were less aggregated and oxidized than LDL from asm+/+/ldlr-/- mice. When tested in vitro, the propensity for aggregation was dependent on the SM level: only LDL from animals on modified diet that have high SM content aggregated when treated with recombinant S-SMase. In conclusion, LDL-SM content and S-SMase activity are up-regulated in mice fed on an atherogenic diet. S-SMase mediates diet-induced changes in LDL ceramide content and aggregation. S-SMase effectiveness in inducing aggregation is dependent on diet-induced enrichment of LDL with SM, possibly through increased hepatic synthesis.
Figures
Similar articles
-
Secretory sphingomyelinase, a product of the acid sphingomyelinase gene, can hydrolyze atherogenic lipoproteins at neutral pH. Implications for atherosclerotic lesion development.J Biol Chem. 1998 Jan 30;273(5):2738-46. doi: 10.1074/jbc.273.5.2738. J Biol Chem. 1998. PMID: 9446580
-
Rabbit aorta and human atherosclerotic lesions hydrolyze the sphingomyelin of retained low-density lipoprotein. Proposed role for arterial-wall sphingomyelinase in subendothelial retention and aggregation of atherogenic lipoproteins.J Clin Invest. 1996 Sep 15;98(6):1455-64. doi: 10.1172/JCI118934. J Clin Invest. 1996. PMID: 8823312 Free PMC article.
-
Increased sphingomyelin content of plasma lipoproteins in apolipoprotein E knockout mice reflects combined production and catabolic defects and enhances reactivity with mammalian sphingomyelinase.J Clin Invest. 1998 Feb 15;101(4):905-12. doi: 10.1172/JCI870. J Clin Invest. 1998. PMID: 9466986 Free PMC article.
-
Sphingomyelinase activity of LDL: a link between atherosclerosis, ceramide, and apoptosis?Trends Cardiovasc Med. 2002 Jan;12(1):37-42. doi: 10.1016/s1050-1738(01)00143-8. Trends Cardiovasc Med. 2002. PMID: 11796243 Review.
-
Secretory sphingomyelinase.Chem Phys Lipids. 1999 Nov;102(1-2):123-30. doi: 10.1016/s0009-3084(99)00080-8. Chem Phys Lipids. 1999. PMID: 11001566 Review.
Cited by
-
Preparation of LDL , Oxidation , Methods of Detection, and Applications in Atherosclerosis Research.Methods Mol Biol. 2022;2419:213-246. doi: 10.1007/978-1-0716-1924-7_13. Methods Mol Biol. 2022. PMID: 35237967
-
Plasma cross-gestational sphingolipidomic analyses reveal potential first trimester biomarkers of preeclampsia.PLoS One. 2017 Apr 6;12(4):e0175118. doi: 10.1371/journal.pone.0175118. eCollection 2017. PLoS One. 2017. PMID: 28384202 Free PMC article.
-
PCSK9 Functions in Atherosclerosis Are Not Limited to Plasmatic LDL-Cholesterol Regulation.Front Cardiovasc Med. 2021 Mar 23;8:639727. doi: 10.3389/fcvm.2021.639727. eCollection 2021. Front Cardiovasc Med. 2021. PMID: 33834043 Free PMC article. Review.
-
Alcohol and high fat induced chronic pancreatitis: TRPV4 antagonist reduces hypersensitivity.Neuroscience. 2015 Dec 17;311:166-79. doi: 10.1016/j.neuroscience.2015.10.028. Epub 2015 Oct 19. Neuroscience. 2015. PMID: 26480812 Free PMC article.
-
ORMDL/serine palmitoyltransferase stoichiometry determines effects of ORMDL3 expression on sphingolipid biosynthesis.J Lipid Res. 2015 Apr;56(4):898-908. doi: 10.1194/jlr.M057539. Epub 2015 Feb 17. J Lipid Res. 2015. PMID: 25691431 Free PMC article.
References
-
- Merrill A. H., Jr, Wang E., Mullins R. E. Kinetics of long-chain (sphingoid) base biosynthesis in intact LM cells: effects of varying the extracellular concentrations of serine and fatty acid precursors of this pathway. Biochemistry. 1988;27:340–345. - PubMed
-
- Merrill A. H., Jr, Lingrell S., Wang E., Nikolova-Karakashian M., Vales T. R., Vance D. E. Sphingolipid biosynthesis de novo by rat hepatocytes in culture. Ceramide and sphingomyelin are associated with, but not required for, very low density lipoprotein secretion. J. Biol. Chem. 1995;270:13834–13841. - PubMed
-
- Tabas I. Secretory sphingomyelinase. Chem. Phys. Lipids. 1999;102:123–130. - PubMed
-
- Oorni K., Posio P., Ala-Korpela M., Jauhiainen M., Kovanen P. T. Sphingomyelinase induces aggregation and fusion of small very low-density lipoprotein and intermediate-density lipoprotein particles and increases their retention to human arterial proteoglycans. Arterioscler. Thromb. Vasc. Biol. 2005;25:1678–1683. - PubMed
-
- Schissel S. L., Jiang X., Tweedie-Hardman J., Jeong T., Camejo E. H., Najib J., Rapp J. H., Williams K. J., Tabas I. Secretory sphingomyelinase, a product of the acid sphingomyelinase gene, can hydrolyze atherogenic lipoproteins at neutral pH. Implications for atherosclerotic lesion development. J. Biol. Chem. 1998;273:2738–2746. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources