

MUT-16 promotes formation of perinuclear mutator foci required for RNA silencing in the C. elegans germline
- PMID: 22713602
- PMCID: PMC3403012
- DOI: 10.1101/gad.193904.112
MUT-16 promotes formation of perinuclear mutator foci required for RNA silencing in the C. elegans germline
Abstract
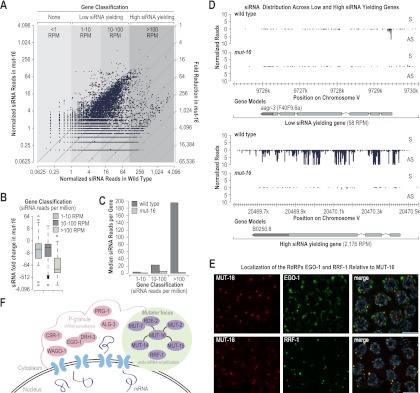
RNA silencing can be initiated by endogenous or exogenously delivered siRNAs. In Caenorhabditis elegans, RNA silencing guided by primary siRNAs is inefficient and therefore requires an siRNA amplification step involving RNA-dependent RNA polymerases (RdRPs). Many factors involved in RNA silencing localize to protein- and RNA-rich nuclear pore-associated P granules in the germline, where they are thought to surveil mRNAs as they exit the nucleus. Mutator class genes are required for siRNA-mediated RNA silencing in both germline and somatic cells, but their specific roles and relationship to other siRNA factors are unclear. Here we show that each of the six mutator proteins localizes to punctate foci at the periphery of germline nuclei. The Mutator foci are adjacent to P granules but are not dependent on core P-granule components or other RNAi pathway factors for their formation or stability. The glutamine/asparagine (Q/N)-rich protein MUT-16 is specifically required for the formation of a protein complex containing the mutator proteins, and in its absence, Mutator foci fail to form at the nuclear periphery. The RdRP RRF-1 colocalizes with MUT-16 at Mutator foci, suggesting a role for Mutator foci in siRNA amplification. Furthermore, we demonstrate that genes that yield high levels of siRNAs, indicative of multiple rounds of siRNA amplification, are disproportionally affected in mut-16 mutants compared with genes that yield low levels of siRNAs. We propose that the mutator proteins and RRF-1 constitute an RNA processing compartment required for siRNA amplification and RNA silencing.
Figures
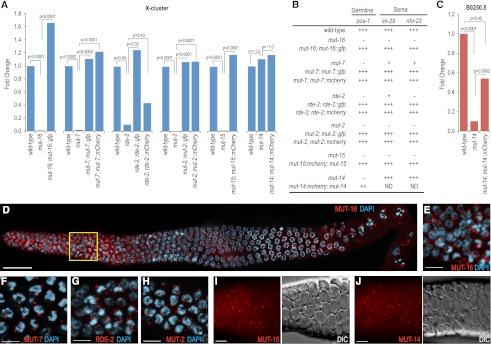
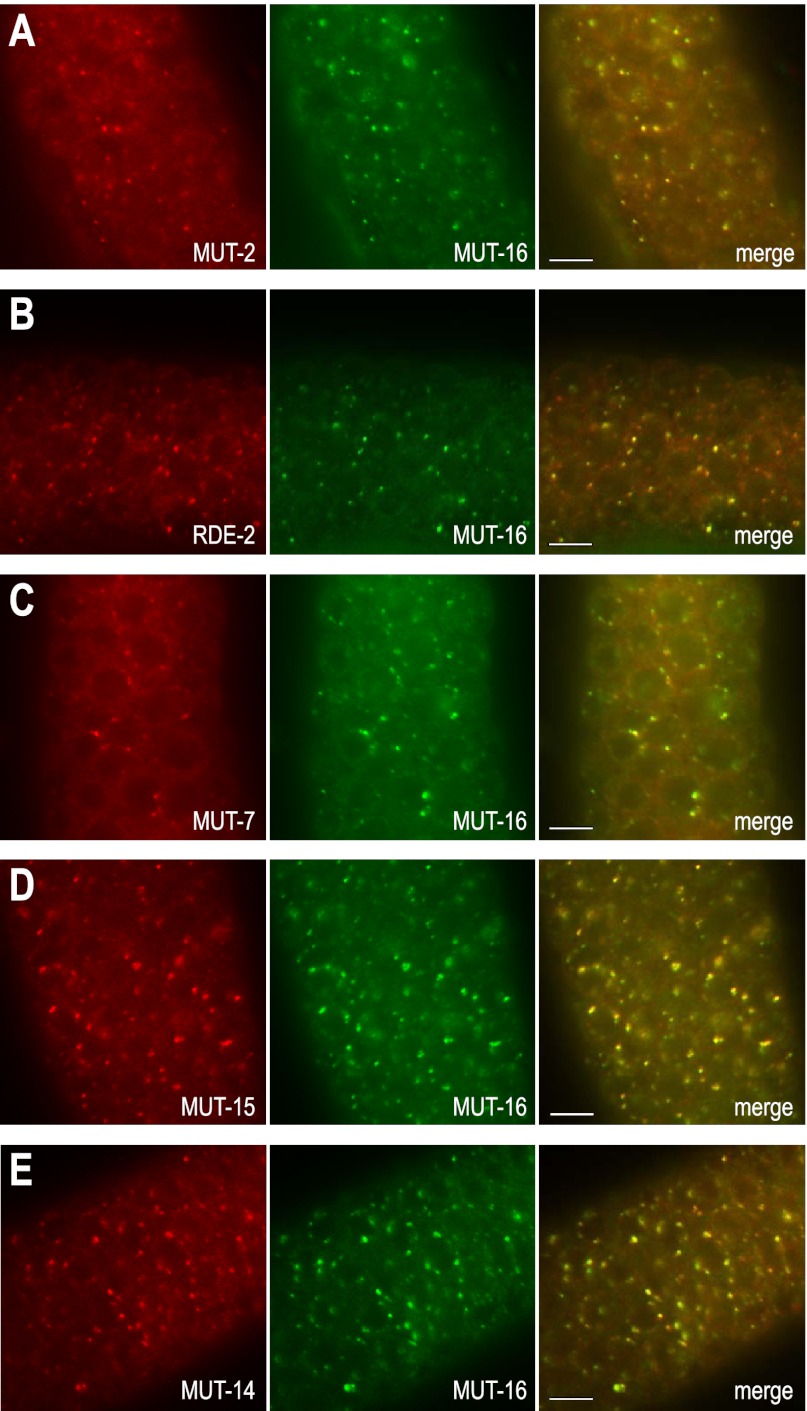
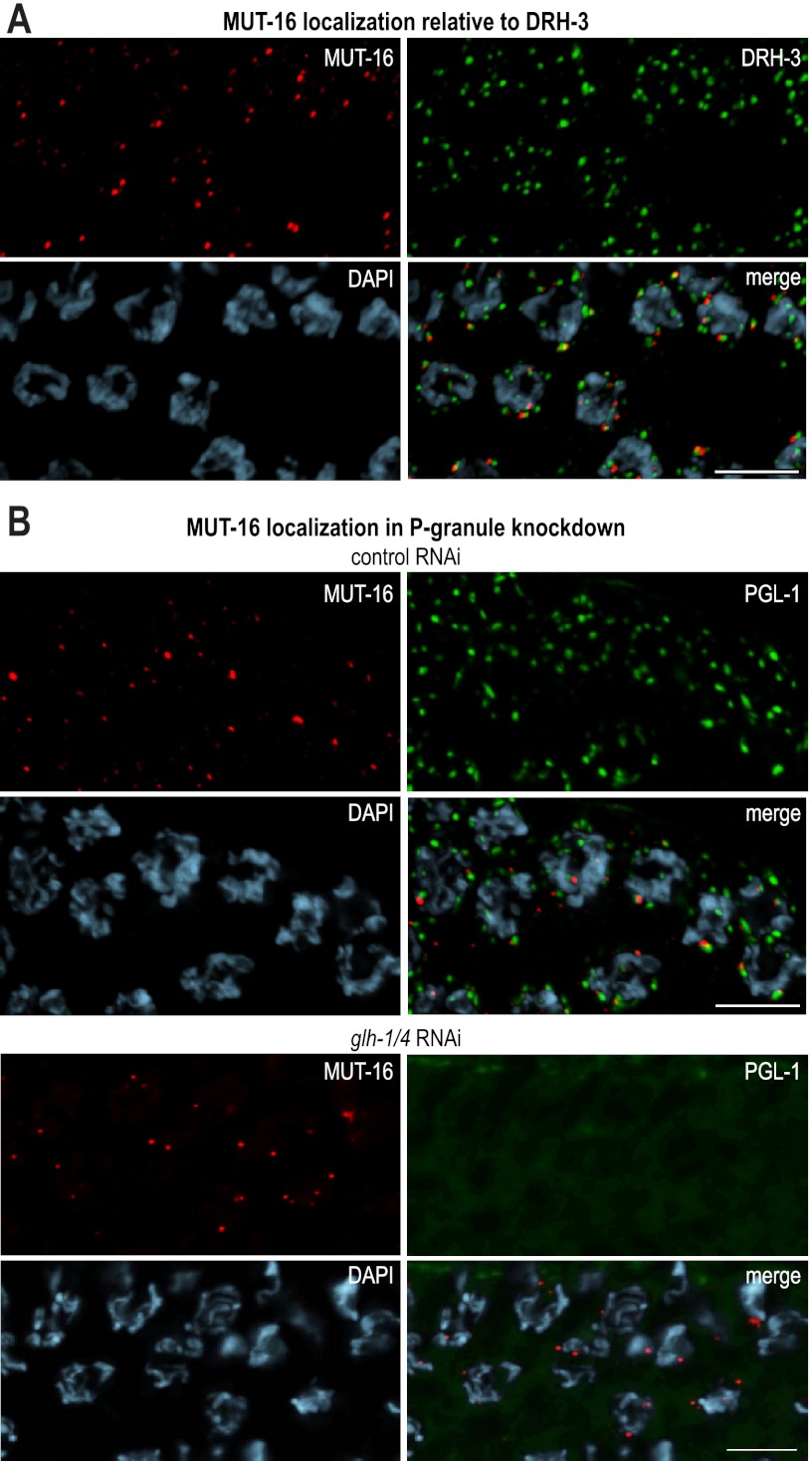

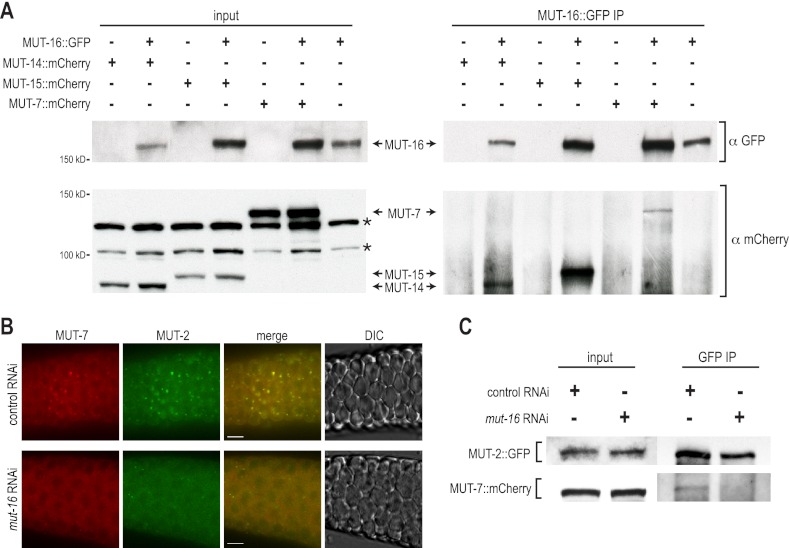
References
-
- Bernstein E, Caudy AA, Hammond SM, Hannon GJ 2001. Role for a bidentate ribonuclease in the initiation step of RNA interference. Nature 409: 363–366 - PubMed
-
- Chen C-CG, Simard MJ, Tabara H, Brownell DR, McCollough JA, Mello CC 2005. A member of the polymerase β nucleotidyltransferase superfamily is required for RNA interference in C. elegans. Curr Biol 15: 378–383 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials