

Mutant p53 aggregates into prion-like amyloid oligomers and fibrils: implications for cancer
- PMID: 22715097
- PMCID: PMC3431633
- DOI: 10.1074/jbc.M112.340638
Mutant p53 aggregates into prion-like amyloid oligomers and fibrils: implications for cancer
Abstract
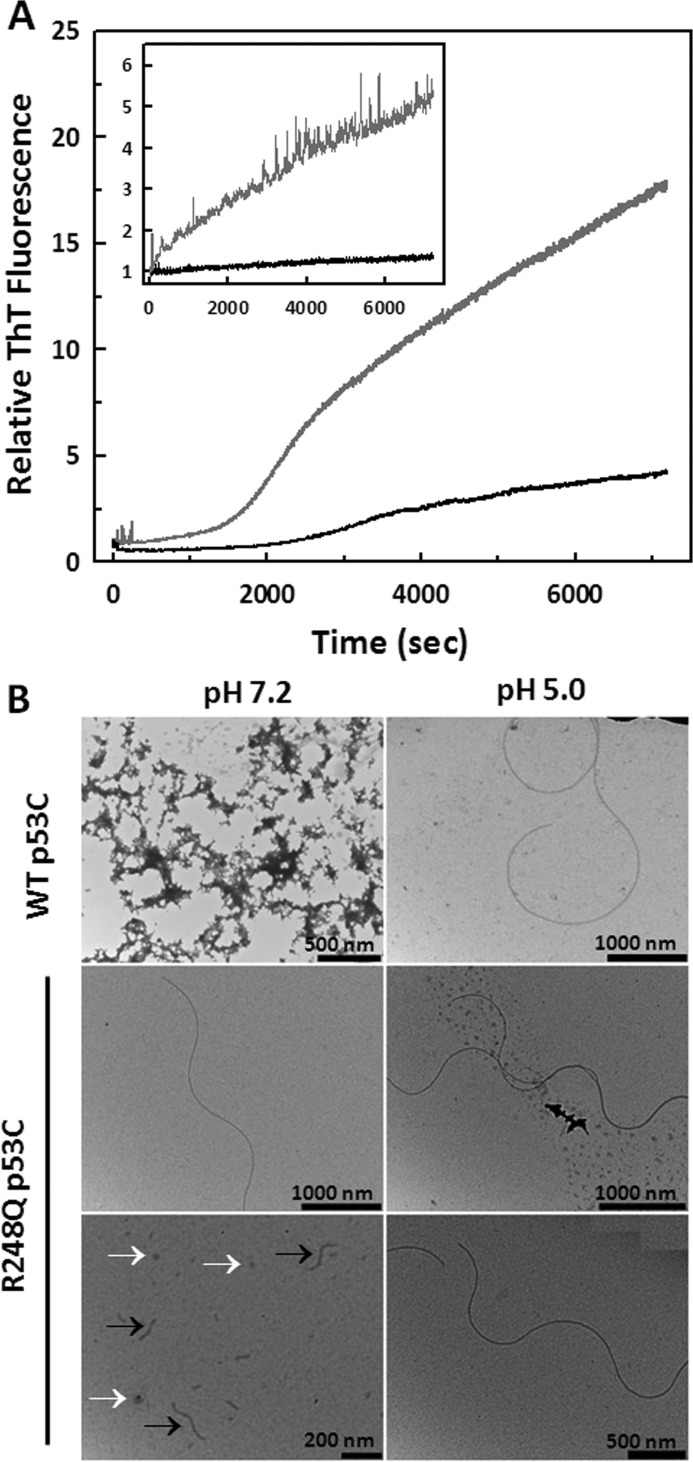
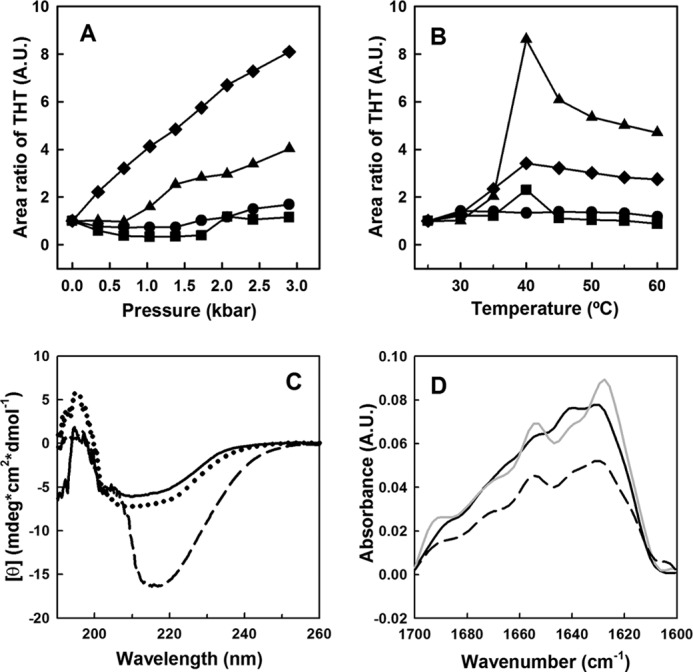
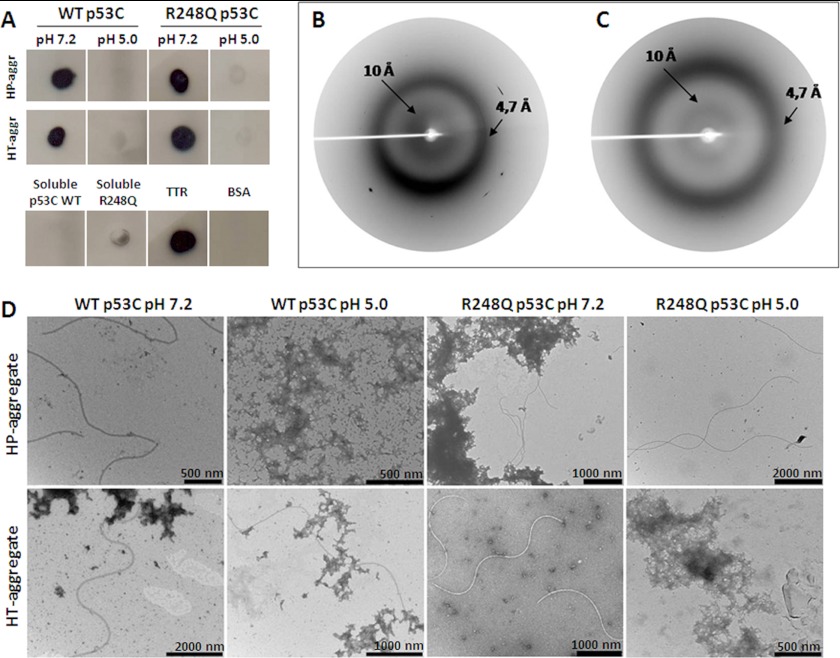
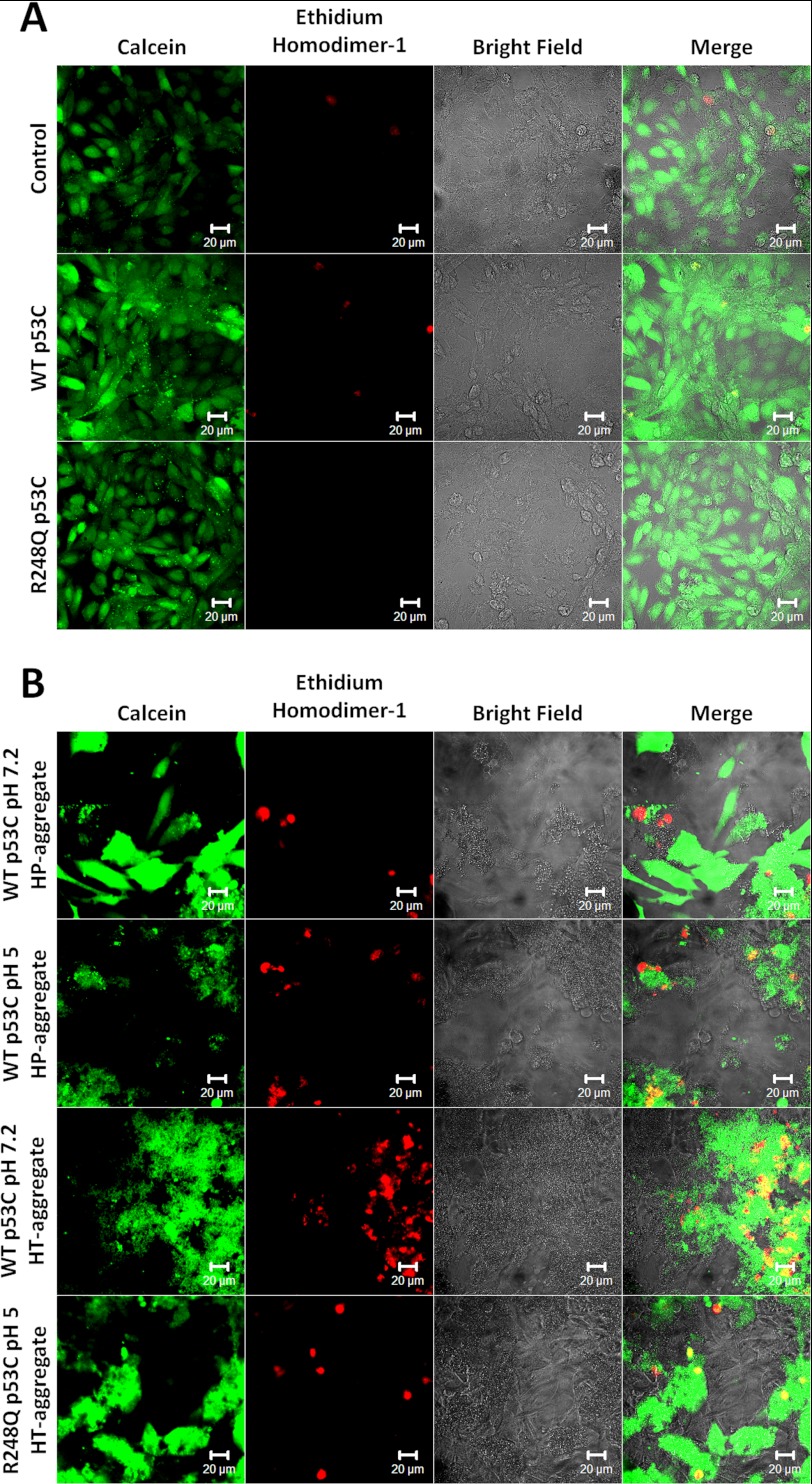
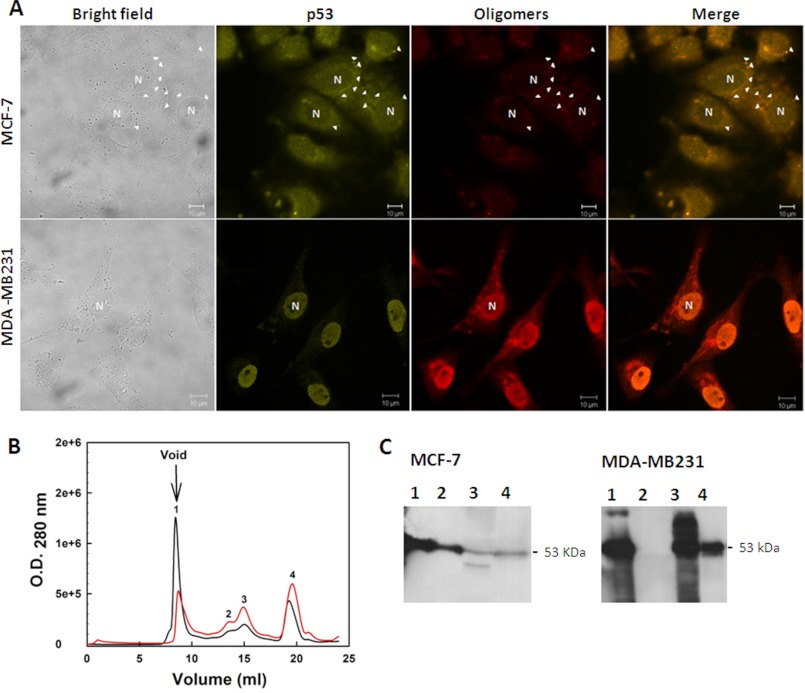
Over 50% of all human cancers lose p53 function. To evaluate the role of aggregation in cancer, we asked whether wild-type (WT) p53 and the hot-spot mutant R248Q could aggregate as amyloids under physiological conditions and whether the mutant could seed aggregation of the wild-type form. The central domains (p53C) of both constructs aggregated into a mixture of oligomers and fibrils. R248Q had a greater tendency to aggregate than WT p53. Full-length p53 aggregated into amyloid-like species that bound thioflavin T. The amyloid nature of the aggregates was demonstrated using x-ray diffraction, electron microscopy, FTIR, dynamic light scattering, cell viabilility assay, and anti-amyloid immunoassay. The x-ray diffraction pattern of the fibrillar aggregates was consistent with the typical conformation of cross β-sheet amyloid fibers with reflexions of 4.7 Å and 10 Å. A seed of R248Q p53C amyloid oligomers and fibrils accelerated the aggregation of WT p53C, a behavior typical of a prion. The R248Q mutant co-localized with amyloid-like species in a breast cancer sample, which further supported its prion-like effect. A tumor cell line containing mutant p53 also revealed massive aggregation of p53 in the nucleus. We conclude that aggregation of p53 into a mixture of oligomers and fibrils sequestrates the native protein into an inactive conformation that is typical of a prionoid. This prion-like behavior of oncogenic p53 mutants provides an explanation for the negative dominance effect and may serve as a potential target for cancer therapy.
Figures





References
-
- Vousden K. H., Lane D. P. (2007) p53 in health and disease. Nat. Rev. Mol. Cell Biol. 8, 275–283 - PubMed
-
- Joerger A. C., Fersht A. R. (2008) Structural biology of the tumor suppressor p53. Annu. Rev. Biochem. 77, 557–582 - PubMed
-
- Ishimaru D., Andrade L. R., Teixeira L. S., Quesado P. A., Maiolino L. M., Lopez P. M., Cordeiro Y., Costa L. T., Heckl W. M., Weissmüller G., Foguel D., Silva J. L. (2003) Fibrillar aggregates of the tumor suppressor p53 core domain. Biochemistry 42, 9022–9027 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
