

Mechanisms and implications of programmed translational frameshifting
- PMID: 22715123
- PMCID: PMC3419312
- DOI: 10.1002/wrna.1126
Mechanisms and implications of programmed translational frameshifting
Abstract
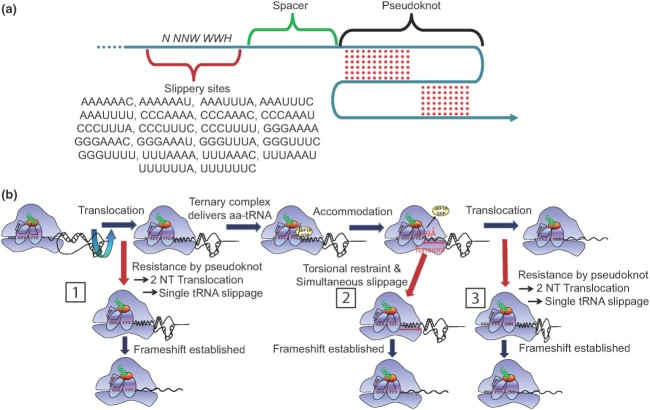
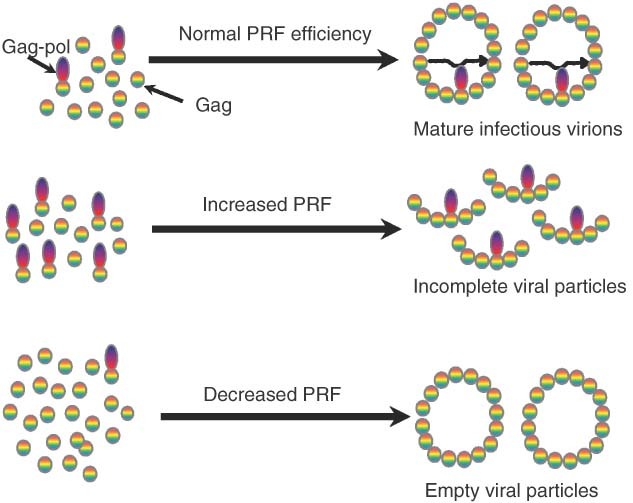
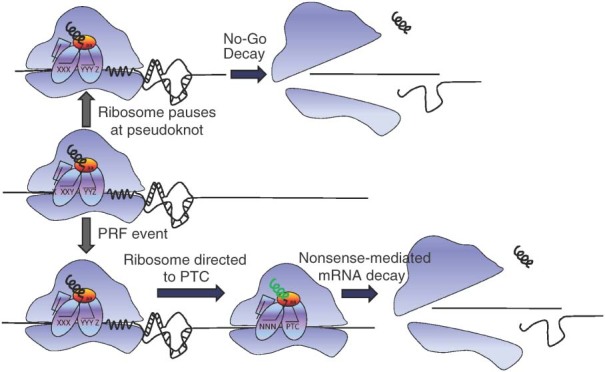
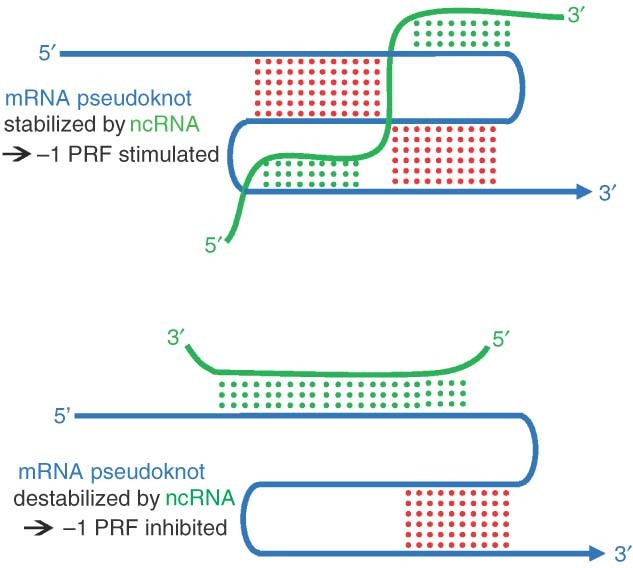
While ribosomes must maintain translational reading frame in order to translate primary genetic information into polypeptides, cis-acting signals located in mRNAs represent higher order information content that can be used to fine-tune gene expression. Classes of signals have been identified that direct a fraction of elongating ribosomes to shift reading frame by one base in the 5' (-1) or 3' (+1) direction. This is called programmed ribosomal frameshifting (PRF). Although mechanisms of PRF differ, a common feature is induction of ribosome pausing, which alters kinetic partitioning rates between in-frame and out-of-frame codons at specific 'slippery' sequences. Many viruses use PRF to ensure synthesis of the correct ratios of virus-encoded proteins required for proper viral particle assembly and maturation, thus identifying PRF as an attractive target for antiviral therapeutics. In contrast, recent studies indicate that PRF signals may primarily function as mRNA destabilizing elements in cellular mRNAs. These studies suggest that PRF may be used to fine-tune gene expression through mRNA decay pathways. The possible regulation of PRF by noncoding RNAs is also discussed.
Copyright © 2012 John Wiley & Sons, Ltd.
Figures
References
-
- Nirenberg M. Historical review: deciphering the genetic code–a personal account. Trends Biochem Sci 2004, 29:46–54. - PubMed
-
- Schmeing TM, Ramakrishnan V. What recent ribosome structures have revealed about the mechanism of translation. Nature 2009, 461:1234–1242. - PubMed
-
- Leung EK, Suslov N, Tuttle N, Sengupta R, Piccirilli JA. The mechanism of peptidyl transfer catalysis by the ribosome. Annu Rev Biochem 2011, 80:527–555. - PubMed
-
- Atkins JF, Gesteland RF. Recoding: Expansion of Decoding RulesEnriches Gene Expression. New York: Springer; 2010..
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
