

A conformational switch controls cell wall-remodelling enzymes required for bacterial cell division
- PMID: 22715947
- PMCID: PMC3418388
- DOI: 10.1111/j.1365-2958.2012.08138.x
A conformational switch controls cell wall-remodelling enzymes required for bacterial cell division
Abstract
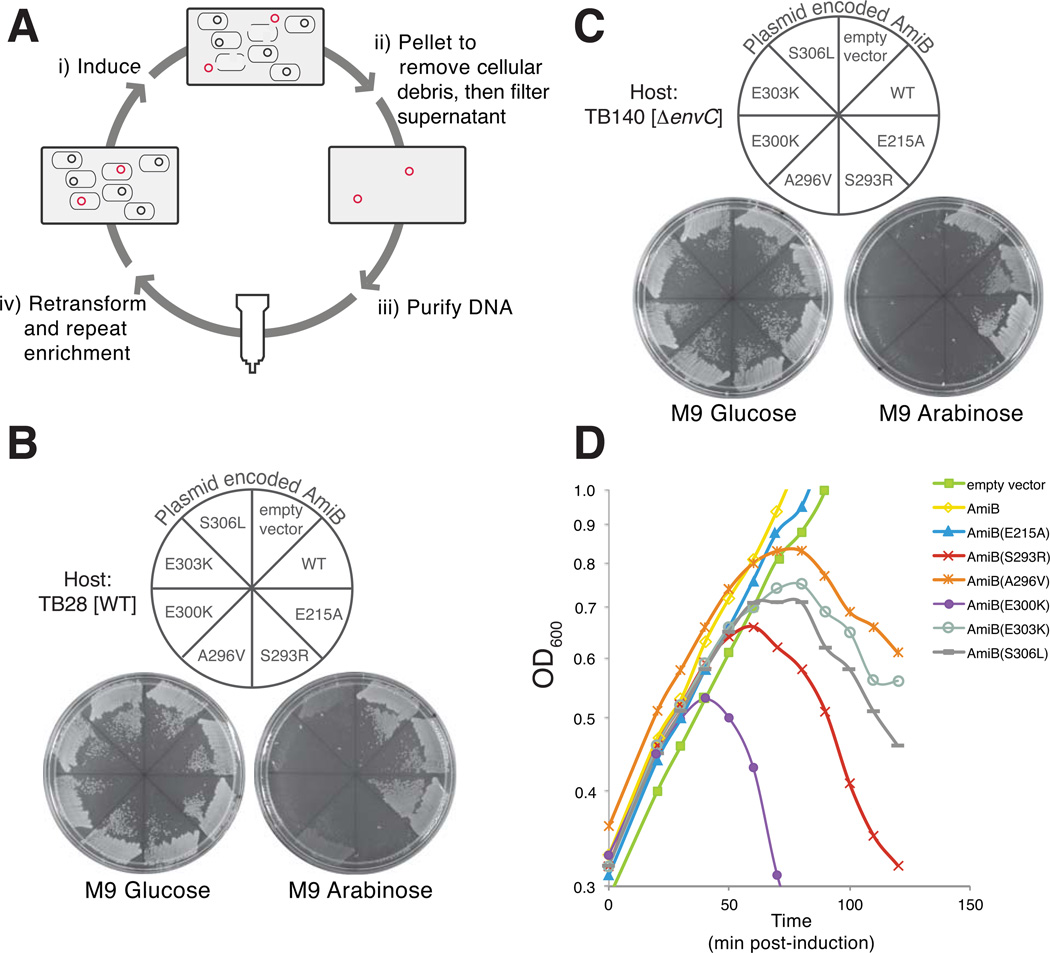
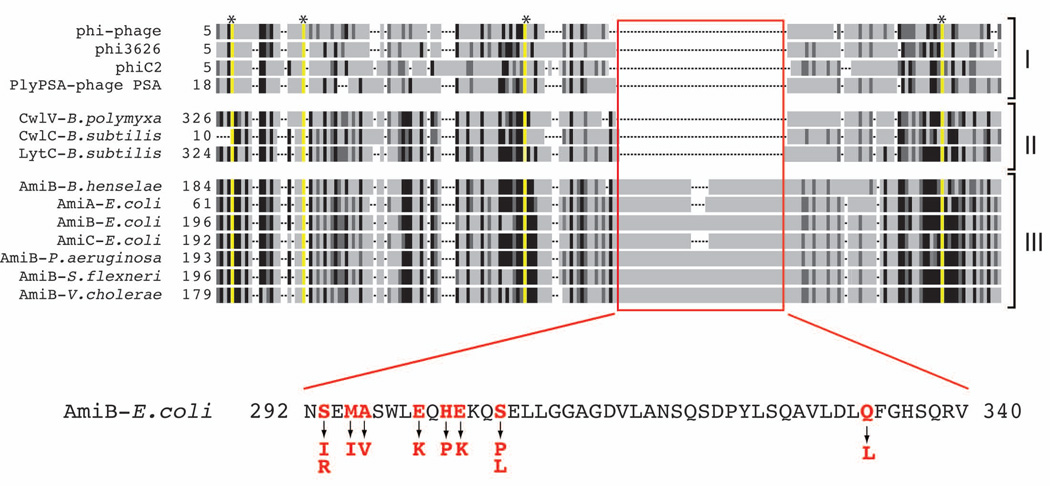
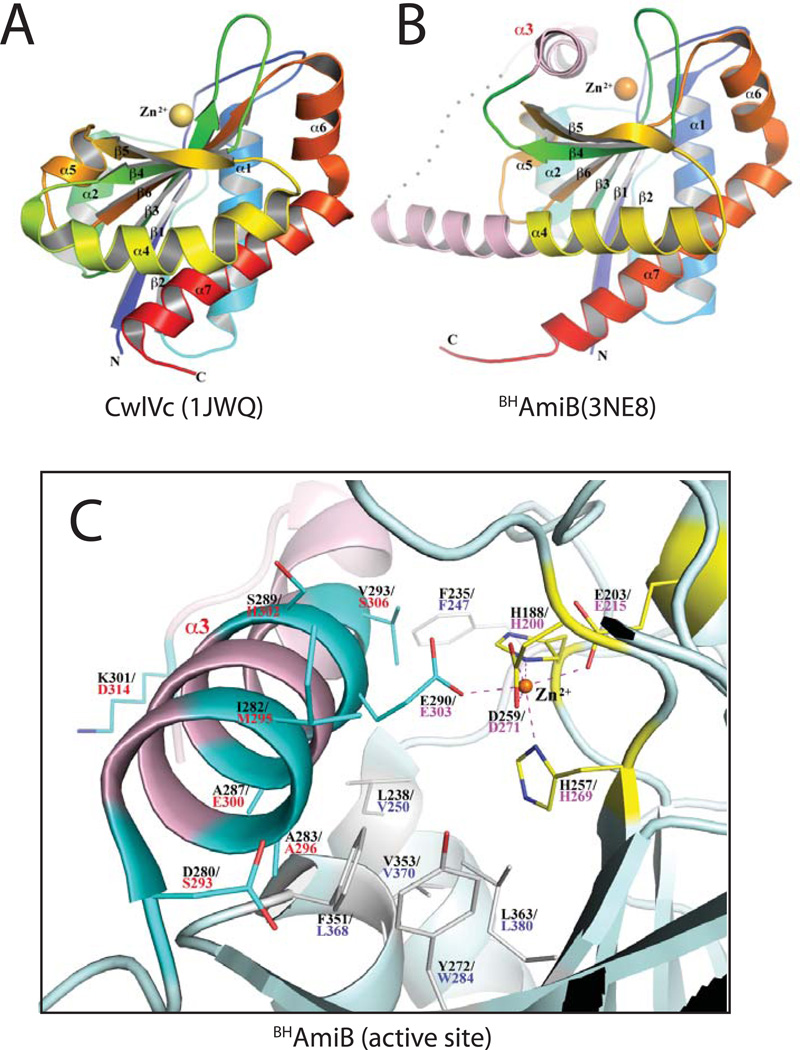
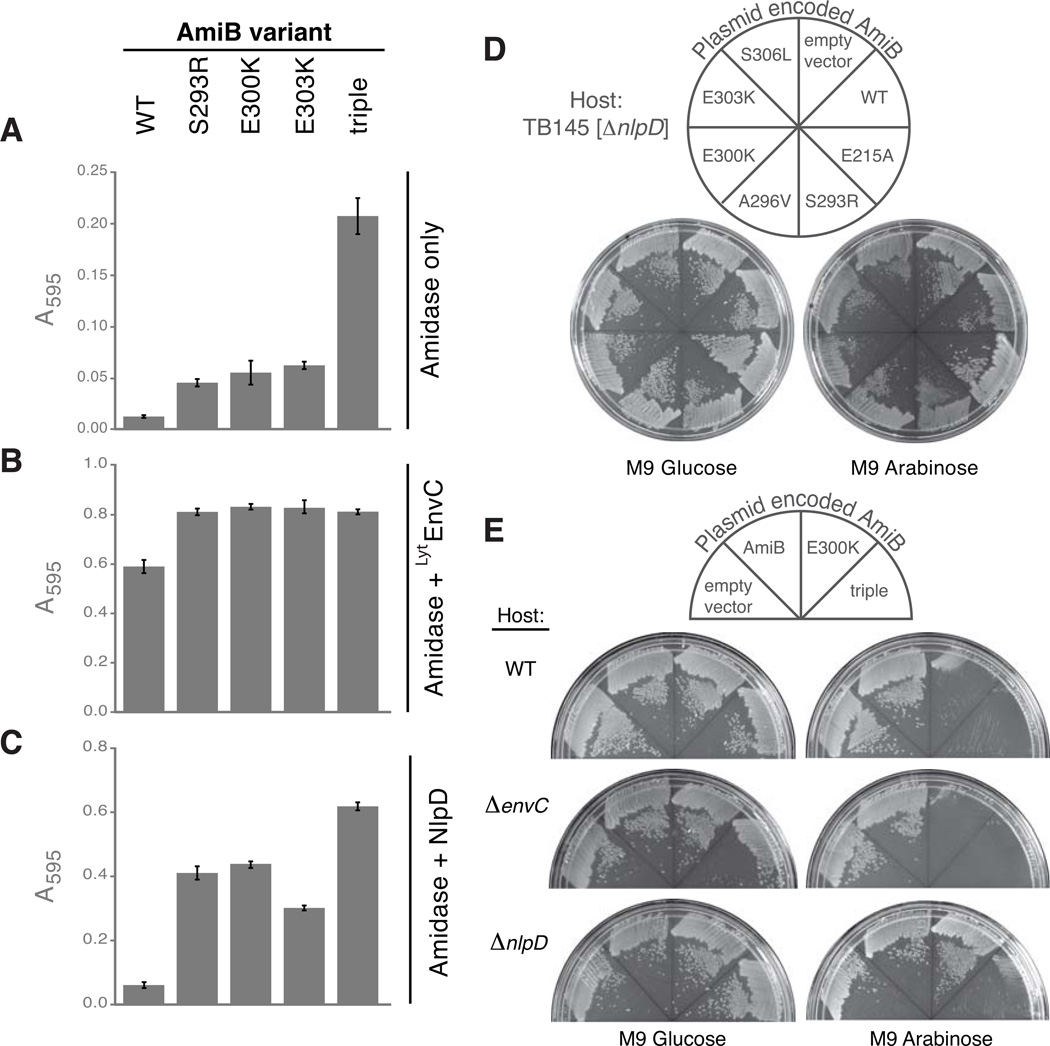
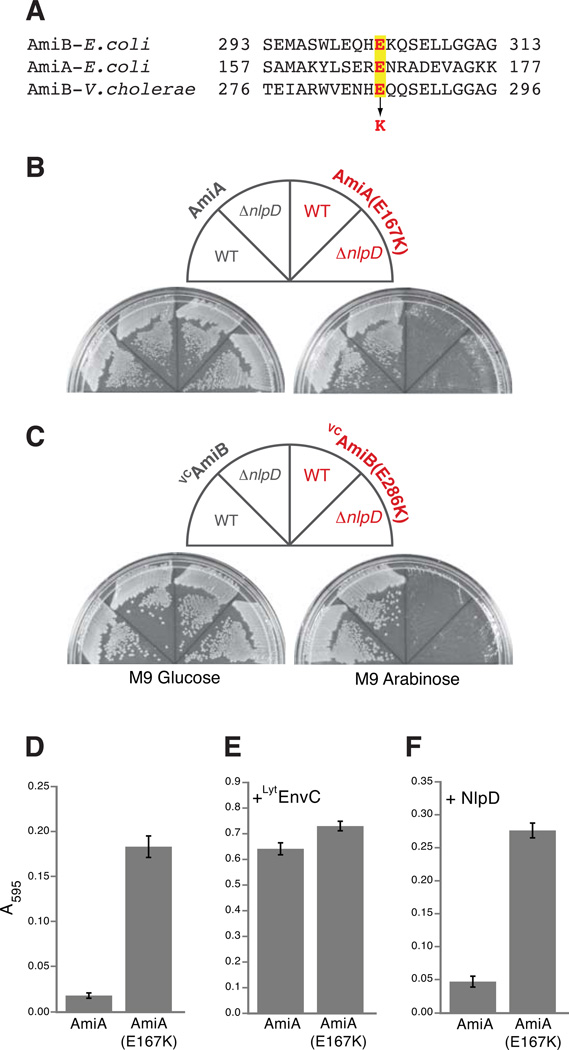
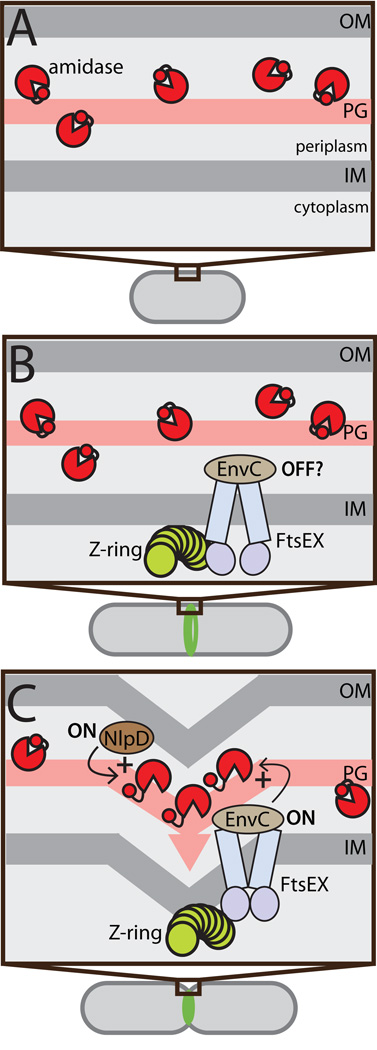
Remodelling of the peptidoglycan (PG) exoskeleton is intimately tied to the growth and division of bacteria. Enzymes that hydrolyse PG are critical for these processes, but their activities must be tightly regulated to prevent the generation of lethal breaches in the PG matrix. Despite their importance, the mechanisms regulating PG hydrolase activity have remained elusive. Here we investigate the control of cell division hydrolases called amidases (AmiA, AmiB and AmiC) required for Escherichia coli cell division. Poorly regulated amiB mutants were isolated encoding lytic AmiB variants with elevated basal PG hydrolase activities in vitro. The structure of an AmiB orthologue was also solved, revealing that the active site of AmiB is occluded by a conserved alpha helix. Strikingly, most of the amino acid substitutions in the lytic AmiB variants mapped to this domain and are predicted to disrupt its interaction with the active site. Our results therefore support a model in which cell separation is stimulated by the reversible relief of amidase autoinhibition governed by conserved subcomplexes within the cytokinetic ring. Analogous conformational control mechanisms are likely to be part of a general strategy used to control PG hydrolases present within multienzyme PG-remodelling machines.
© 2012 Blackwell Publishing Ltd.
Figures
References
-
- Bi EF, Lutkenhaus J. FtsZ ring structure associated with division in Escherichia coli. Nature. 1991;354:161–164. - PubMed
-
- Böth D, Schneider G, Schnell R. Peptidoglycan Remodeling in Mycobacterium tuberculosis: Comparison of Structures and Catalytic Activities of RipA and RipB. J Mol Biol. 2011;413:247–260. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
