

CaMKII regulation in information processing and storage
- PMID: 22717267
- PMCID: PMC3461103
- DOI: 10.1016/j.tins.2012.05.003
CaMKII regulation in information processing and storage
Abstract
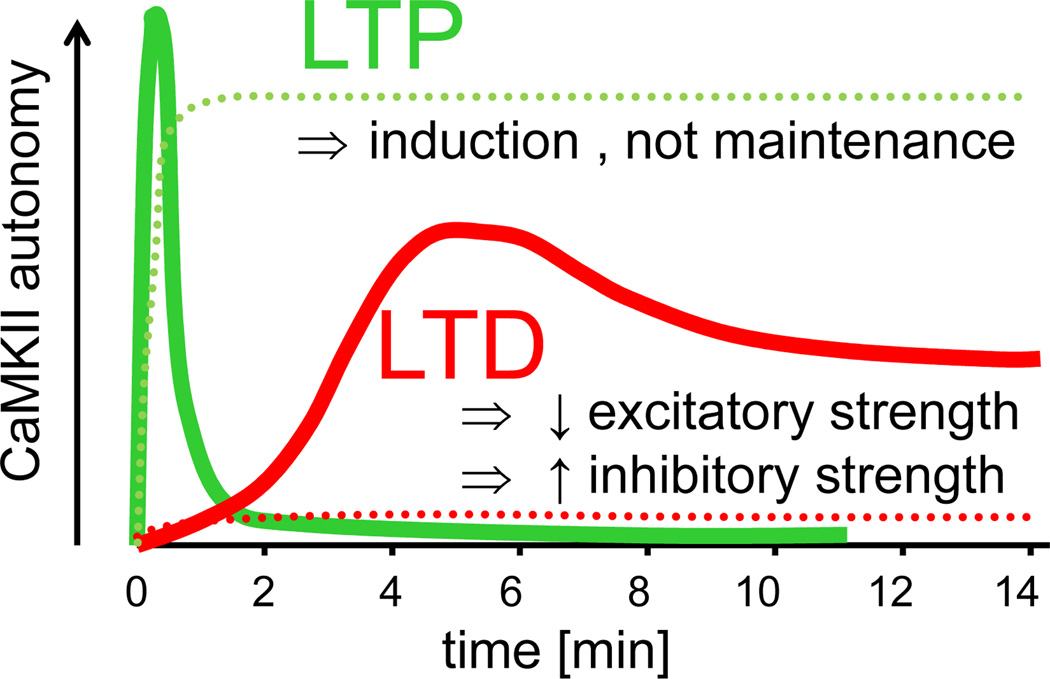
The Ca(2+)/Calmodulin(CaM)-dependent protein kinase II (CaMKII) is activated by Ca(2+)/CaM, but becomes partially autonomous (Ca(2+)-independent) upon autophosphorylation at T286. This hallmark feature of CaMKII regulation provides a form of molecular memory and is indeed important in long-term potentiation (LTP) of excitatory synapse strength and memory formation. However, emerging evidence supports a direct role in information processing, while storage of synaptic information may instead be mediated by regulated interaction of CaMKII with the NMDA receptor (NMDAR) complex. These and other CaMKII regulation mechanisms are discussed here in the context of the kinase structure and their impact on postsynaptic functions. Recent findings also implicate CaMKII in long-term depression (LTD), as well as functional roles at inhibitory synapses, lending renewed emphasis on better understanding the spatiotemporal control of CaMKII regulation.
Copyright © 2012 Elsevier Ltd. All rights reserved.
Figures
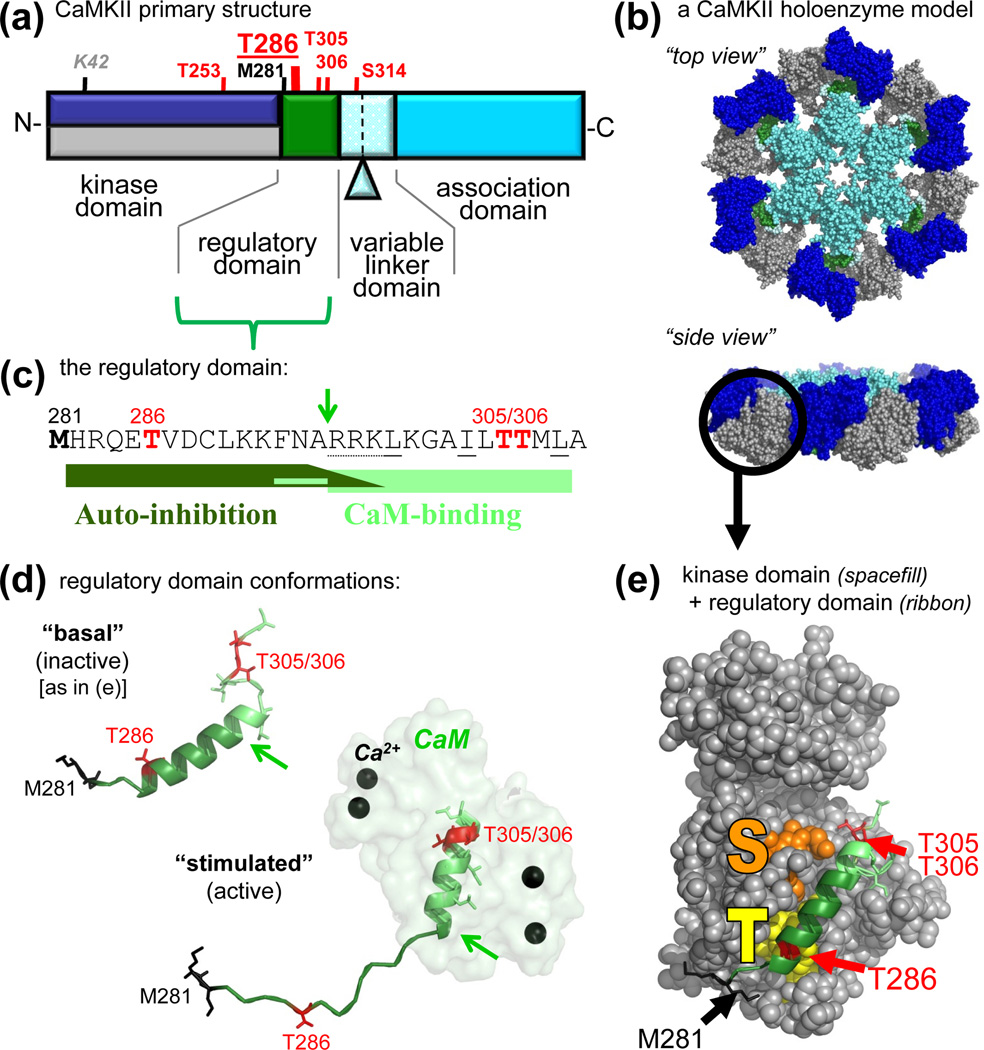
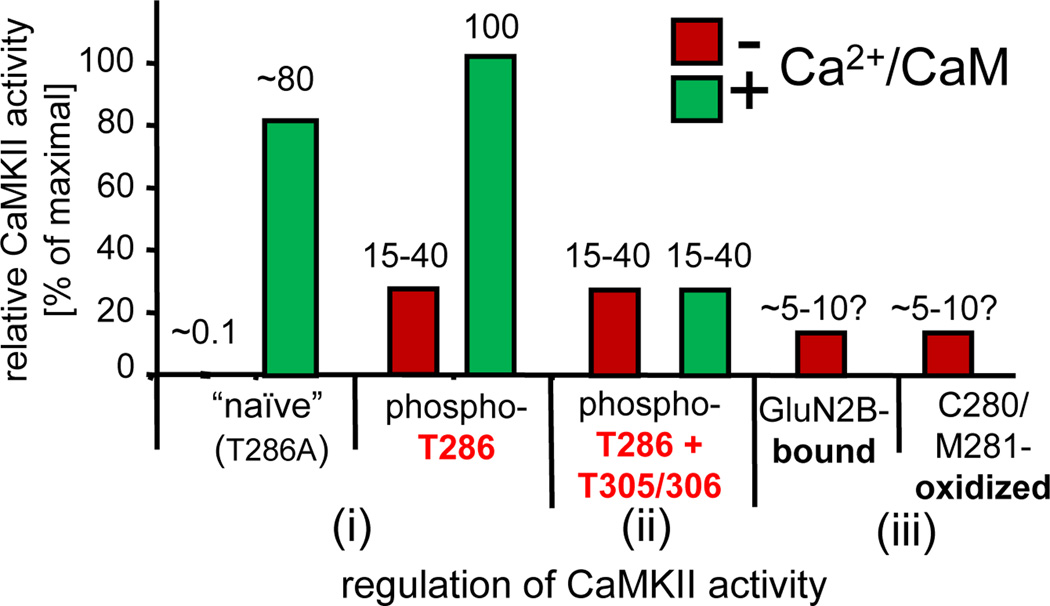
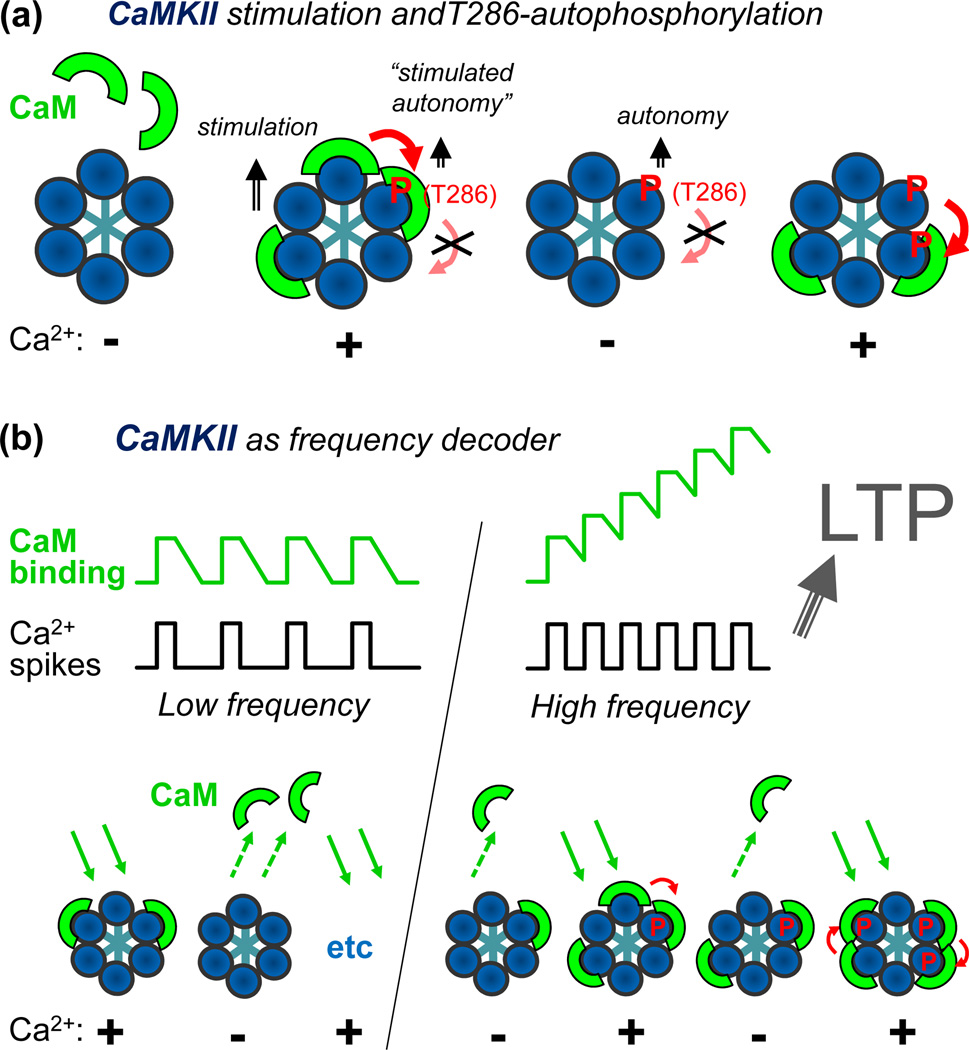
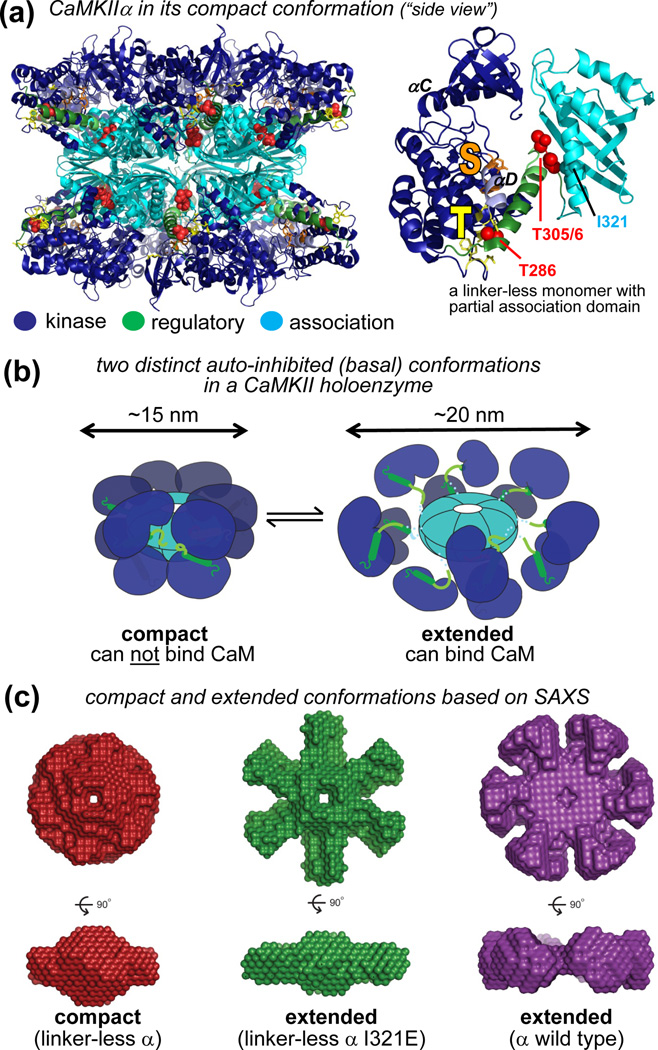
References
-
- Schulman H, Greengard P. Stimulation of brain membrane protein phosphorylation by calcium and an endogenous heat-stable protein. Nature. 1978;271:478–479. - PubMed
-
- Hudmon A, Schulman H. Neuronal CA2+/calmodulin-dependent protein kinase II: the role of structure and autoregulation in cellular function. Annu Rev Biochem. 2002;71:473–510. - PubMed
-
- Malinow R, et al. Inhibition of postsynaptic PKC or CaMKII blocks induction but not expression of LTP. Science. 1989;245:862–866. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
