

Molecular chaperones and stress-inducible protein-sorting factors coordinate the spatiotemporal distribution of protein aggregates
- PMID: 22718905
- PMCID: PMC3418301
- DOI: 10.1091/mbc.E12-03-0194
Molecular chaperones and stress-inducible protein-sorting factors coordinate the spatiotemporal distribution of protein aggregates
Abstract
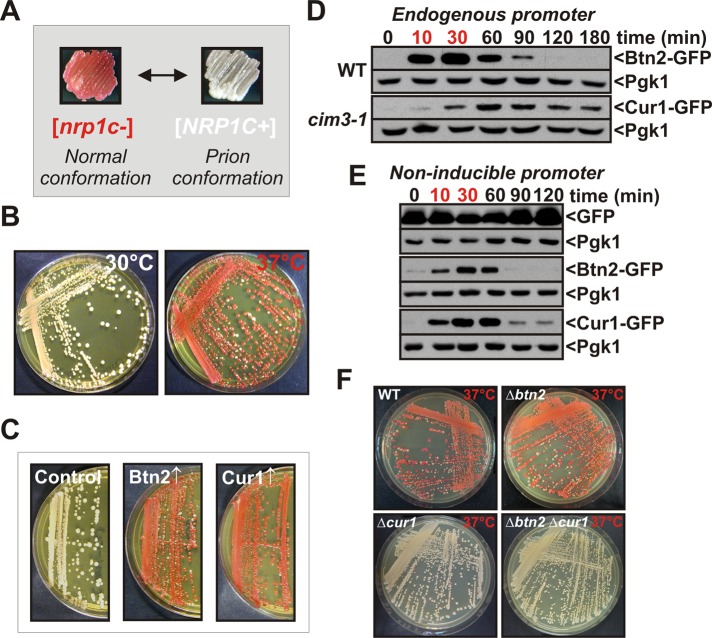
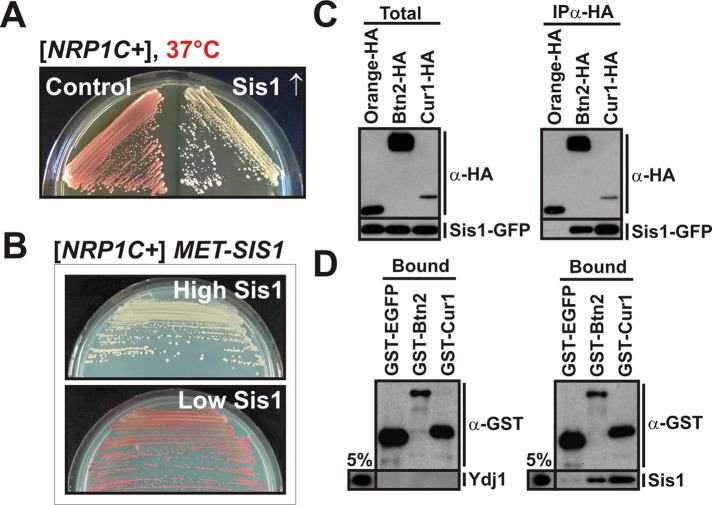
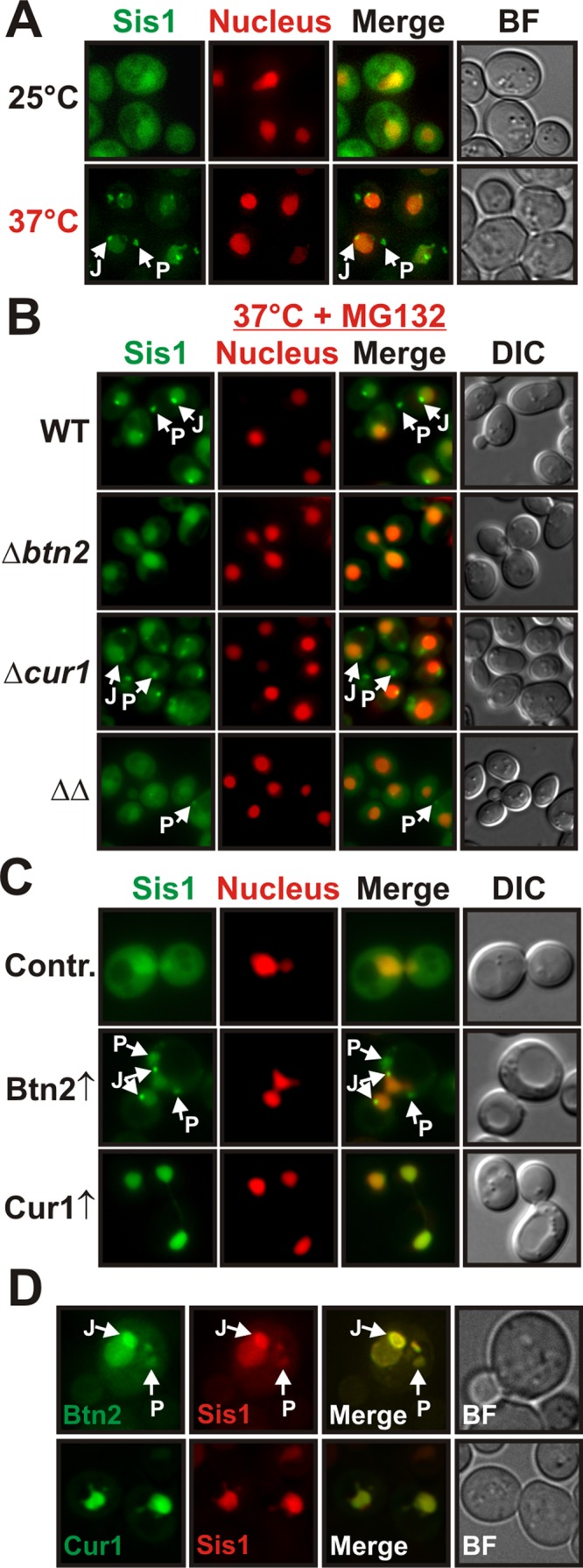
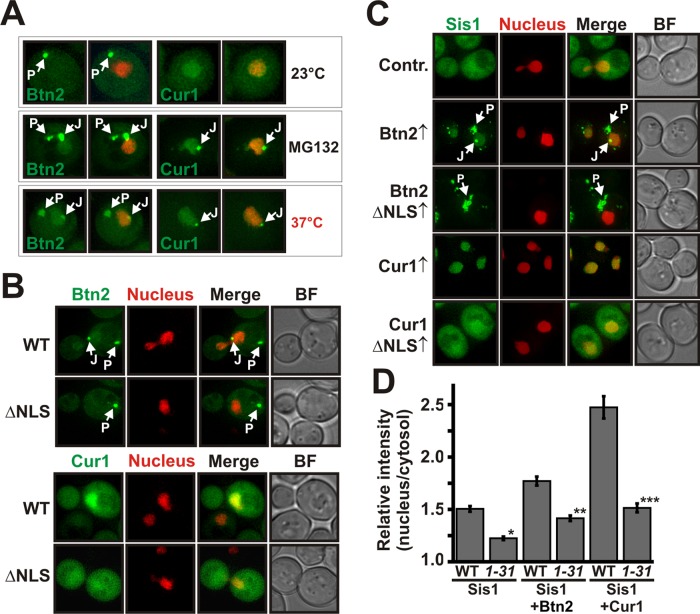
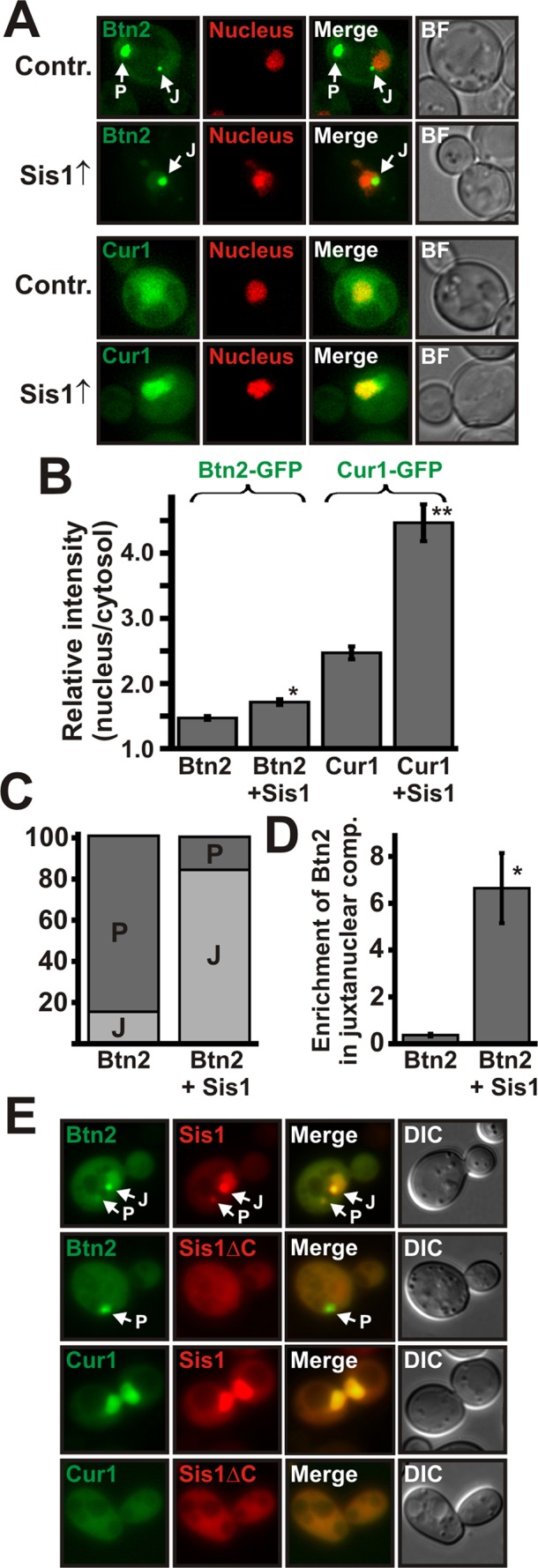
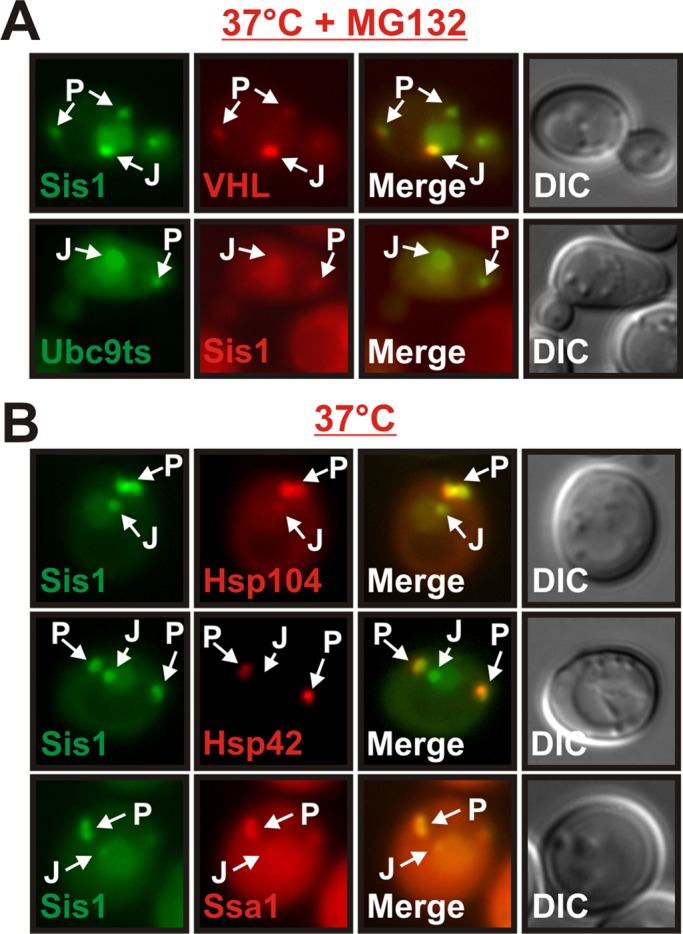
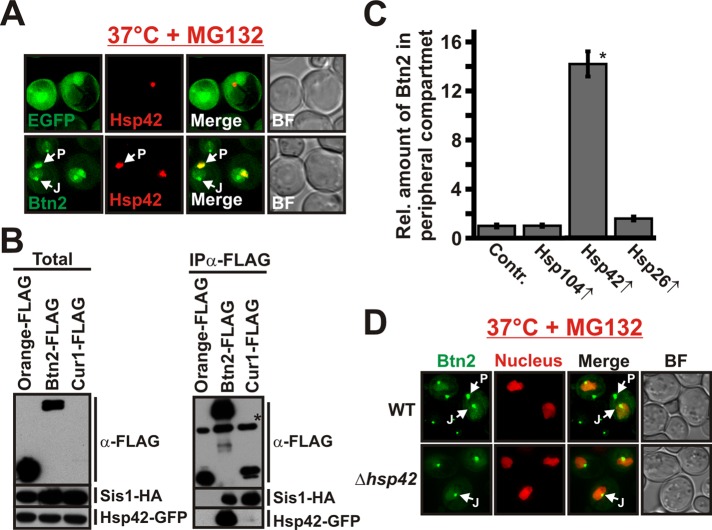
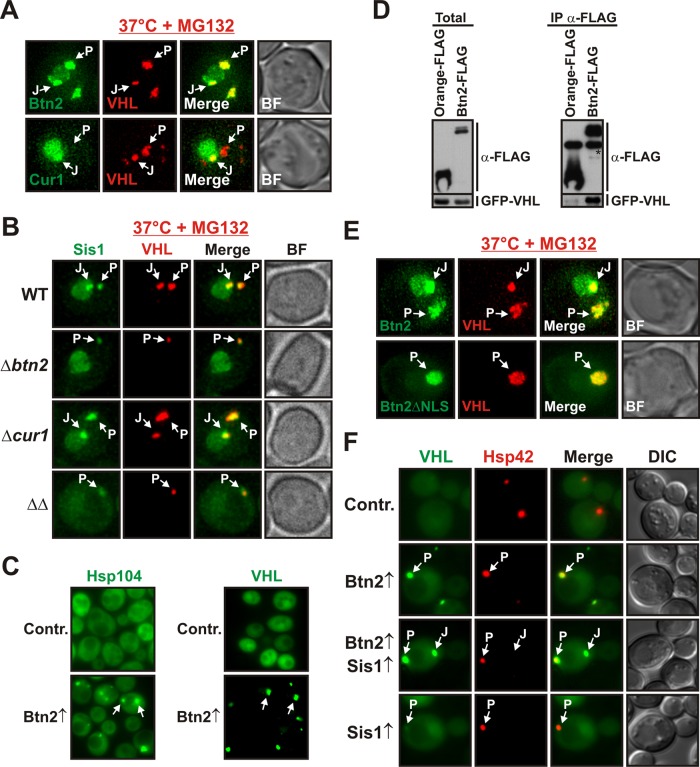
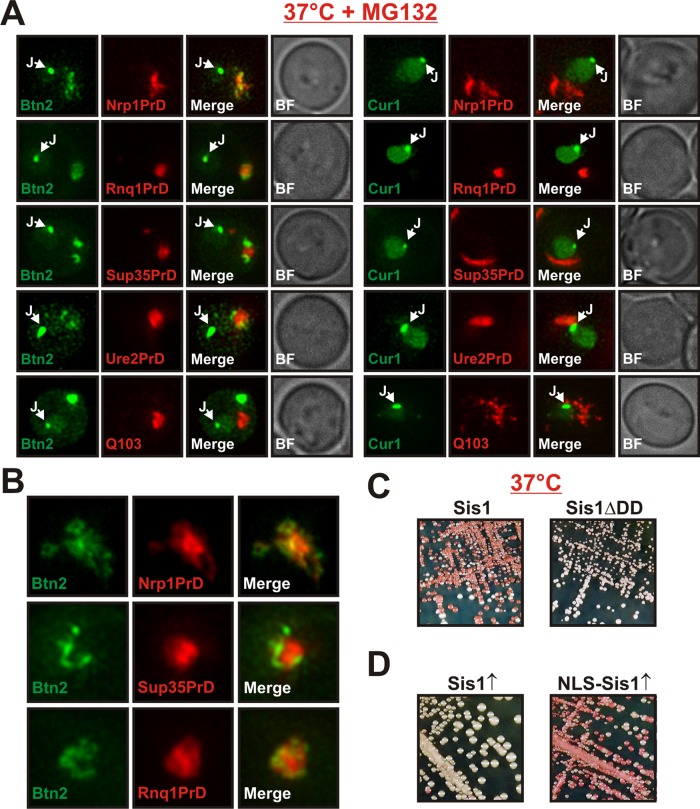
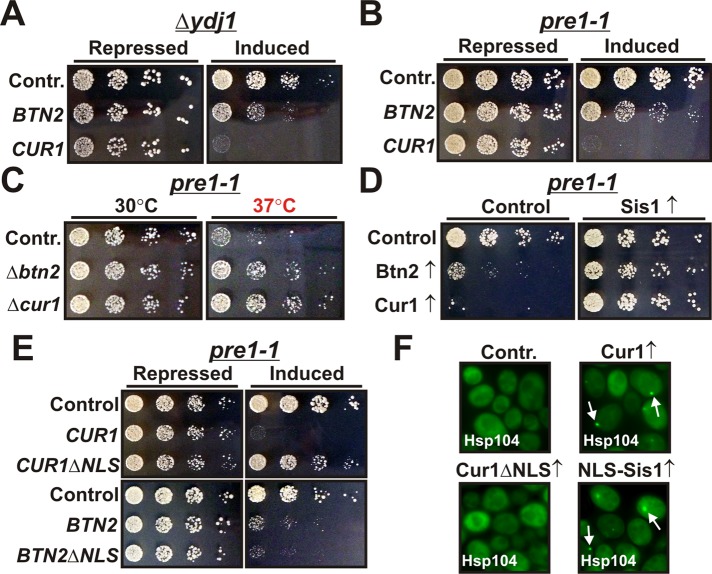
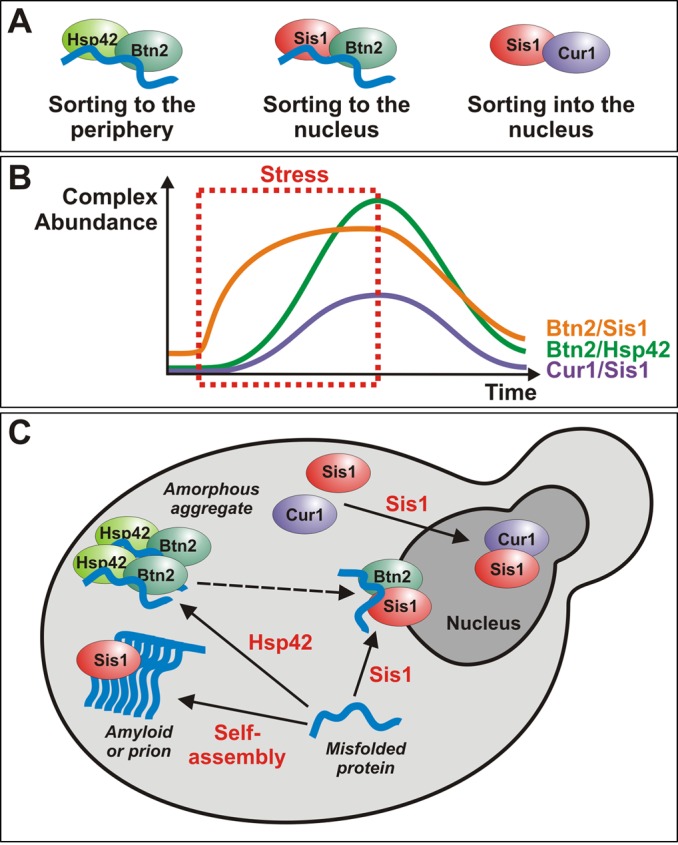
Acute stress causes a rapid redistribution of protein quality control components and aggregation-prone proteins to diverse subcellular compartments. How these remarkable changes come about is not well understood. Using a phenotypic reporter for a synthetic yeast prion, we identified two protein-sorting factors of the Hook family, termed Btn2 and Cur1, as key regulators of spatial protein quality control in Saccharomyces cerevisiae. Btn2 and Cur1 are undetectable under normal growth conditions but accumulate in stressed cells due to increased gene expression and reduced proteasomal turnover. Newly synthesized Btn2 can associate with the small heat shock protein Hsp42 to promote the sorting of misfolded proteins to a peripheral protein deposition site. Alternatively, Btn2 can bind to the chaperone Sis1 to facilitate the targeting of misfolded proteins to a juxtanuclear compartment. Protein redistribution by Btn2 is accompanied by a gradual depletion of Sis1 from the cytosol, which is mediated by the sorting factor Cur1. On the basis of these findings, we propose a dynamic model that explains the subcellular distribution of misfolded proteins as a function of the cytosolic concentrations of molecular chaperones and protein-sorting factors. Our model suggests that protein aggregation is not a haphazard process but rather an orchestrated cellular response that adjusts the flux of misfolded proteins to the capacities of the protein quality control system.
Figures
References
-
- Alberti S, Halfmann R, Lindquist S. Biochemical, cell biological, and genetic assays to analyze amyloid and prion aggregation in yeast. Methods Enzymol. 2010;470:709–734. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
