

Regulation of p21/CIP1/WAF-1 mediated cell-cycle arrest by RNase L and tristetraprolin, and involvement of AU-rich elements
- PMID: 22718976
- PMCID: PMC3439922
- DOI: 10.1093/nar/gks545
Regulation of p21/CIP1/WAF-1 mediated cell-cycle arrest by RNase L and tristetraprolin, and involvement of AU-rich elements
Abstract
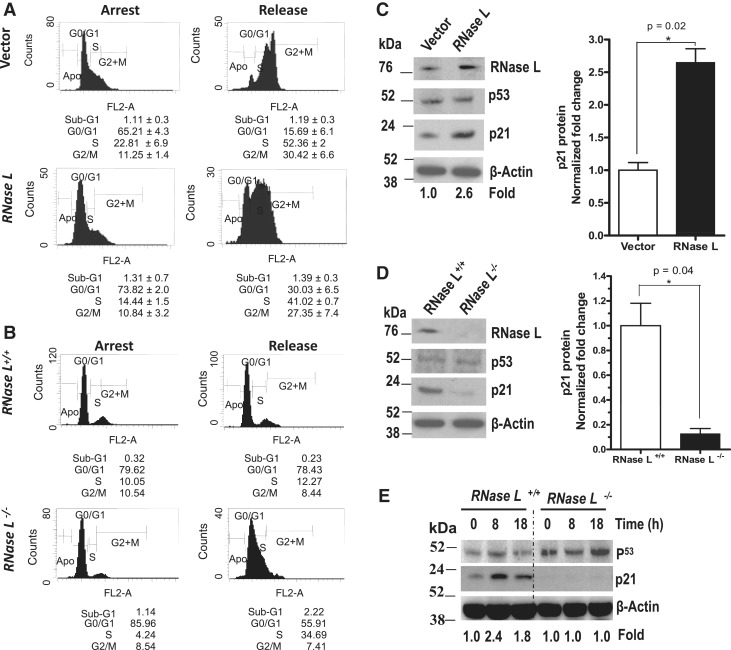
The p21(Cip1/WAF1) plays an important role in cell-cycle arrest. Here, we find that RNase L regulates p21-mediated G(1) growth arrest in AU-rich elements-dependent manner. We found a significant loss of p21 mRNA expression in RNASEL(-/-) MEFs and that the overexpression of RNase L in HeLa cells induces p21 mRNA expression. The p21 mRNA half-life significantly changes as a result of RNase L modulation, indicating a post-transcriptional effect. Indeed, we found that RNase L promotes tristetraprolin (TTP/ZFP36) mRNA decay. This activity was not seen with dimerization- and nuclease-deficient RNase L mutants. Deficiency in TTP led to increases in p21 mRNA and protein. With induced ablation of RNase L, TTP mRNA and protein expressions were higher, while p21 expression became reduced. We further establish that TTP, but not C124R TTP mutant, binds to, and accelerates the decay of p21 mRNA. The p21 mRNA half-life was prolonged in TTP(-/-) MEFs. The TTP regulation of p21 mRNA decay required functional AU-rich elements. Thus, we demonstrate a novel mechanism of regulating G(1) growth arrest by an RNase L-TTP-p21 axis.
Figures






References
-
- Khabar KS, Dhalla M, Siddiqui Y, Zhou A, Al-Ahdal MN, Der SD, Silverman RH, Williams BR. Effect of deficiency of the double-stranded RNA-dependent protein kinase, PKR, on antiviral resistance in the presence or absence of ribonuclease L: HSV-1 replication is particularly sensitive to deficiency of the major IFN-mediated enzymes. J. Interferon Cytokine Res. 2000;20:653–659. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
