

IGF-1 restores visual cortex plasticity in adult life by reducing local GABA levels
- PMID: 22720172
- PMCID: PMC3375096
- DOI: 10.1155/2012/250421
IGF-1 restores visual cortex plasticity in adult life by reducing local GABA levels
Abstract
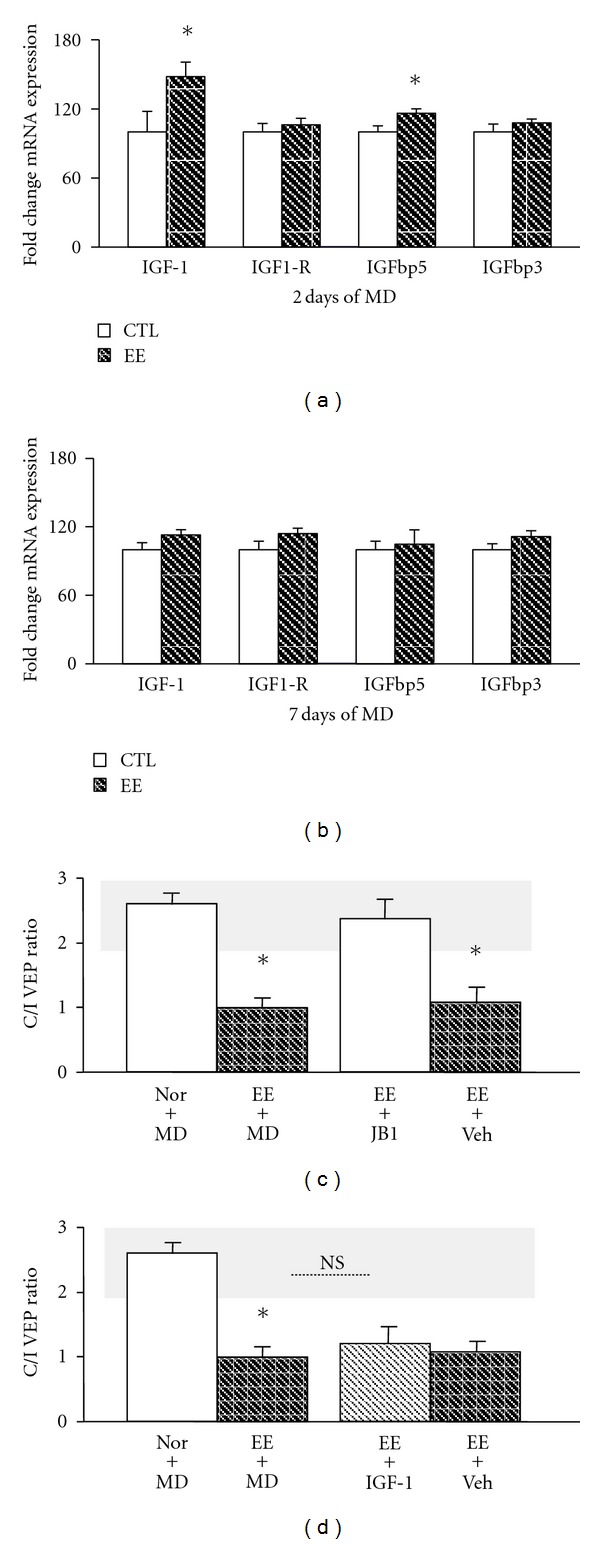
The central nervous system architecture is markedly modified by sensory experience during early life, but a decline of plasticity occurs with age. Recent studies have challenged this dogma providing evidence that both pharmacological treatments and paradigms based on the manipulation of environmental stimulation levels can be successfully employed as strategies for enhancing plasticity in the adult nervous system. Insulin-like growth factor 1 (IGF-1) is a peptide implicated in prenatal and postnatal phases of brain development such as neurogenesis, neuronal differentiation, synaptogenesis, and experience-dependent plasticity. Here, using the visual system as a paradigmatic model, we report that IGF-1 reactivates neural plasticity in the adult brain. Exogenous administration of IGF-1 in the adult visual cortex, indeed, restores the susceptibility of cortical neurons to monocular deprivation and promotes the recovery of normal visual functions in adult amblyopic animals. These effects were accompanied by a marked reduction of intracortical GABA levels. Moreover, we show that a transitory increase of IGF-1 expression is associated to the plasticity reinstatement induced by environmental enrichment (EE) and that blocking IGF-1 action by means of the IGF-1 receptor antagonist JB1 prevents EE effects on plasticity processes.
Figures


References
-
- Katz LC, Shatz CJ. Synaptic activity and the construction of cortical circuits. Science. 1996;274(5290):1133–1138. - PubMed
-
- Berardi N, Pizzorusso T, Maffei L. Critical periods during sensory development. Current Opinion in Neurobiology. 2000;10(1):138–145. - PubMed
-
- Hensch TK. Critical period plasticity in local cortical circuits. Nature Reviews Neuroscience. 2005;6(11):877–888. - PubMed
-
- Wiesel T, Hubel DH. Single-cell responses in striate cortex of kittens deprived of vision. Journal of Neurophysiology. 1963;26:1003–1017. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
