

Cortical GABAergic interneurons in cross-modal plasticity following early blindness
- PMID: 22720175
- PMCID: PMC3377178
- DOI: 10.1155/2012/590725
Cortical GABAergic interneurons in cross-modal plasticity following early blindness
Abstract
Early loss of a given sensory input in mammals causes anatomical and functional modifications in the brain via a process called cross-modal plasticity. In the past four decades, several animal models have illuminated our understanding of the biological substrates involved in cross-modal plasticity. Progressively, studies are now starting to emphasise on cell-specific mechanisms that may be responsible for this intermodal sensory plasticity. Inhibitory interneurons expressing γ-aminobutyric acid (GABA) play an important role in maintaining the appropriate dynamic range of cortical excitation, in critical periods of developmental plasticity, in receptive field refinement, and in treatment of sensory information reaching the cerebral cortex. The diverse interneuron population is very sensitive to sensory experience during development. GABAergic neurons are therefore well suited to act as a gate for mediating cross-modal plasticity. This paper attempts to highlight the links between early sensory deprivation, cortical GABAergic interneuron alterations, and cross-modal plasticity, discuss its implications, and further provide insights for future research in the field.
Figures
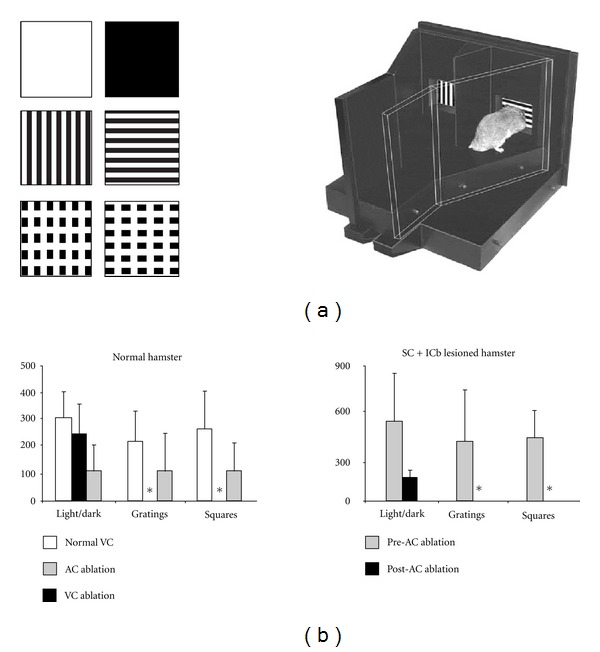
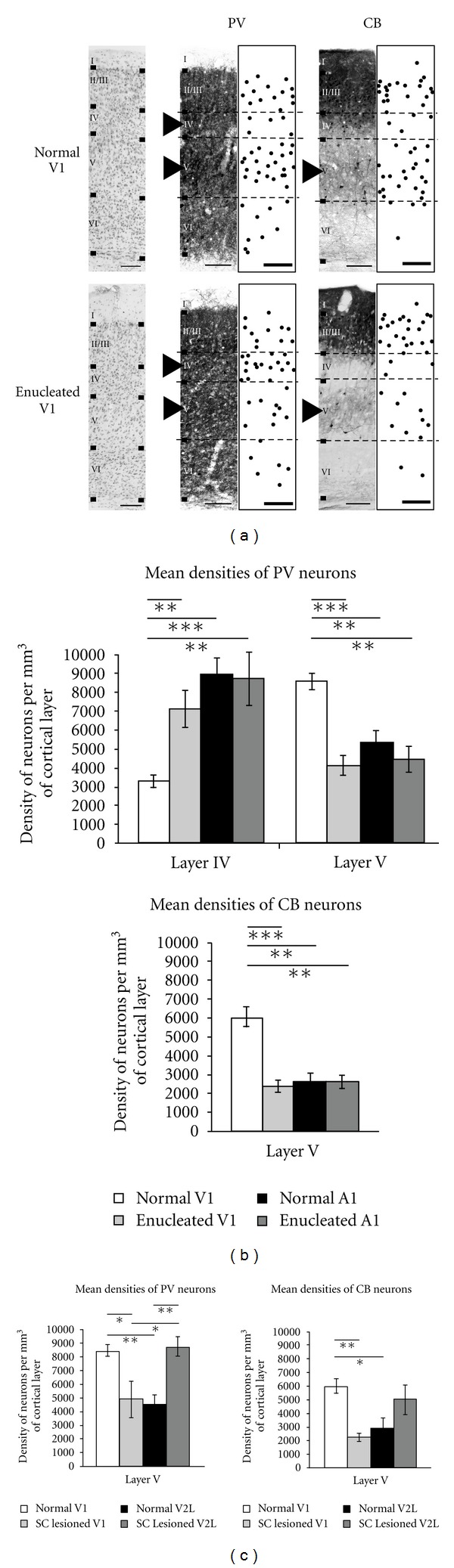
Similar articles
-
Interneuron epigenomes during the critical period of cortical plasticity: Implications for schizophrenia.Neurobiol Learn Mem. 2015 Oct;124:104-10. doi: 10.1016/j.nlm.2015.03.005. Epub 2015 Apr 4. Neurobiol Learn Mem. 2015. PMID: 25849095 Free PMC article. Review.
-
Transient and localized optogenetic activation of somatostatin-interneurons in mouse visual cortex abolishes long-term cortical plasticity due to vision loss.Brain Struct Funct. 2018 Jun;223(5):2073-2095. doi: 10.1007/s00429-018-1611-7. Epub 2018 Jan 25. Brain Struct Funct. 2018. PMID: 29372324 Free PMC article.
-
Parvalbumin-Positive Interneurons Regulate Cortical Sensory Plasticity in Adulthood and Development Through Shared Mechanisms.Front Neural Circuits. 2022 May 6;16:886629. doi: 10.3389/fncir.2022.886629. eCollection 2022. Front Neural Circuits. 2022. PMID: 35601529 Free PMC article. Review.
-
GABA through the ages: regulation of cortical function and plasticity by inhibitory interneurons.Neural Plast. 2012;2012:892784. doi: 10.1155/2012/892784. Epub 2012 Jun 26. Neural Plast. 2012. PMID: 22792496 Free PMC article. Review.
-
Human brain plasticity: evidence from sensory deprivation and altered language experience.Prog Brain Res. 2002;138:177-88. doi: 10.1016/S0079-6123(02)38078-6. Prog Brain Res. 2002. PMID: 12432770 Review.
Cited by
-
Ototoxicity-related changes in GABA immunolabeling within the rat inferior colliculus.Hear Res. 2024 Oct;452:109106. doi: 10.1016/j.heares.2024.109106. Epub 2024 Aug 21. Hear Res. 2024. PMID: 39181061
-
Pain perception is increased in congenital but not late onset blindness.PLoS One. 2014 Sep 22;9(9):e107281. doi: 10.1371/journal.pone.0107281. eCollection 2014. PLoS One. 2014. PMID: 25244529 Free PMC article.
-
Excitatory Crossmodal Input to a Widespread Population of Primary Sensory Cortical Neurons.Neurosci Bull. 2022 Oct;38(10):1139-1152. doi: 10.1007/s12264-022-00855-4. Epub 2022 Apr 16. Neurosci Bull. 2022. PMID: 35429324 Free PMC article.
-
Growth and Behavior of Congenitally Anophthalmic Lee-Sung Pigs.Comp Med. 2019 May 1;69(3):212-220. doi: 10.30802/AALAS-CM-18-000095. Epub 2019 Jun 6. Comp Med. 2019. PMID: 31171049 Free PMC article.
-
Metabolic changes in the visual cortex of binocular blindness macaque monkeys: a proton magnetic resonance spectroscopy study.PLoS One. 2013 Nov 5;8(11):e80073. doi: 10.1371/journal.pone.0080073. eCollection 2013. PLoS One. 2013. PMID: 24224037 Free PMC article.
References
-
- Pallas SL. Intrinsic and extrinsic factors that shape neocortical specification. Trends in Neurosciences. 2001;24(7):417–423. - PubMed
-
- Majewska AK, Sur M. Plasticity and specificity of cortical processing networks. Trends in Neurosciences. 2006;29(6):323–329. - PubMed
-
- O’Leary DD, Chou SJ, Sahara S. Area patterning of the mammalian cortex. Neuron. 2007;56(2):252–269. - PubMed
-
- Ptito M, Desgent S. Sensory input-based adaptation and brain architecture. In: Baltes P, Reuter-Lorenz P, Rosler F, editors. Lifespan Development and the Brain. The Perspective of Biocultural Co-constructivism. Cambridge, UK: Cambridge University Press; 2006. pp. 111–134.
-
- Krubitzer L. The magnificent compromise: cortical field evolution in mammals. Neuron. 2007;56(2):201–208. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
