

Medial prefrontal cortex neuronal activation and synaptic alterations after stress-induced reinstatement of palatable food seeking: a study using c-fos-GFP transgenic female rats
- PMID: 22723688
- PMCID: PMC3389493
- DOI: 10.1523/JNEUROSCI.5895-11.2012
Medial prefrontal cortex neuronal activation and synaptic alterations after stress-induced reinstatement of palatable food seeking: a study using c-fos-GFP transgenic female rats
Abstract
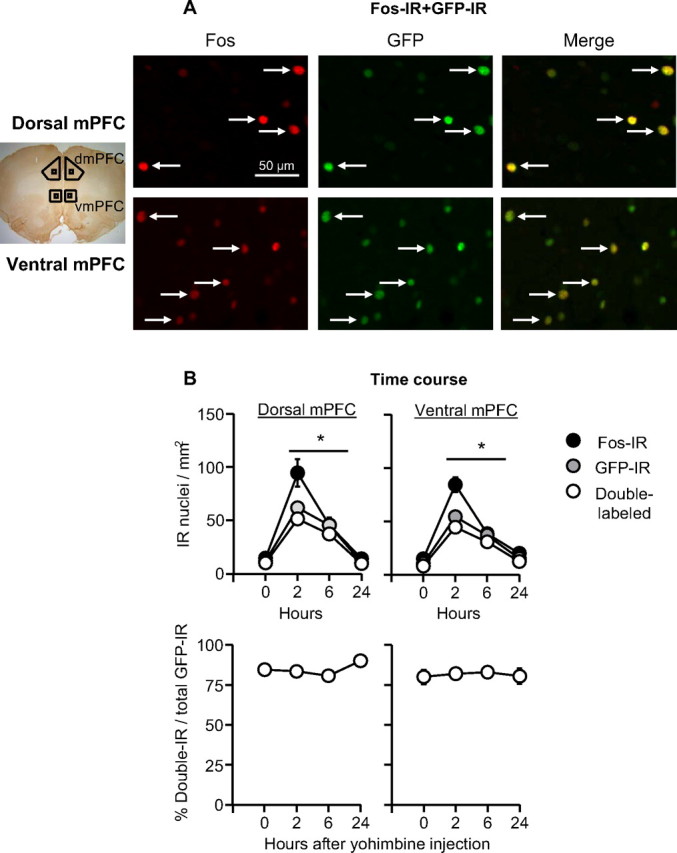
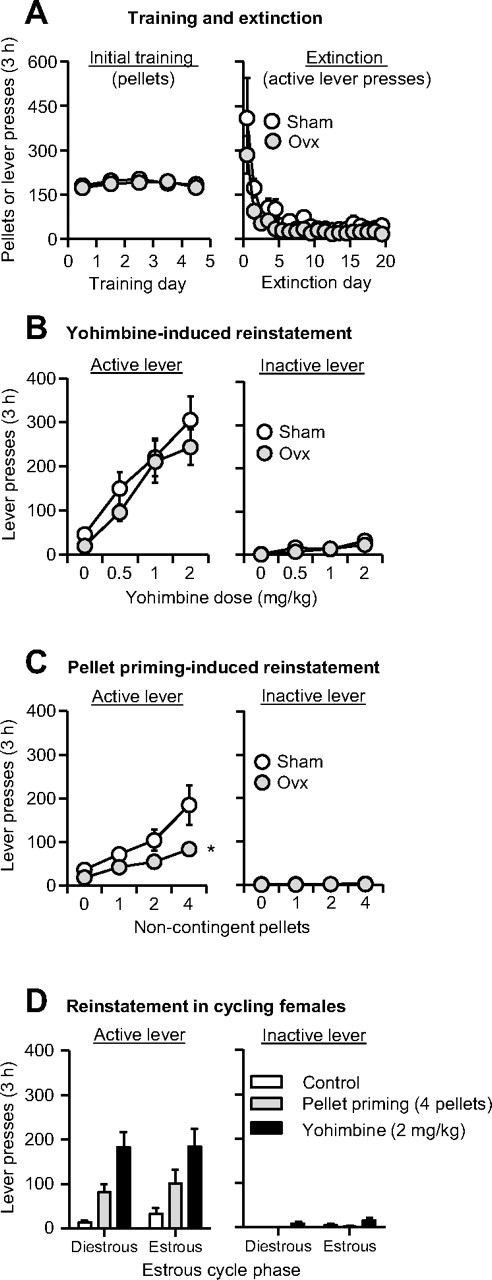
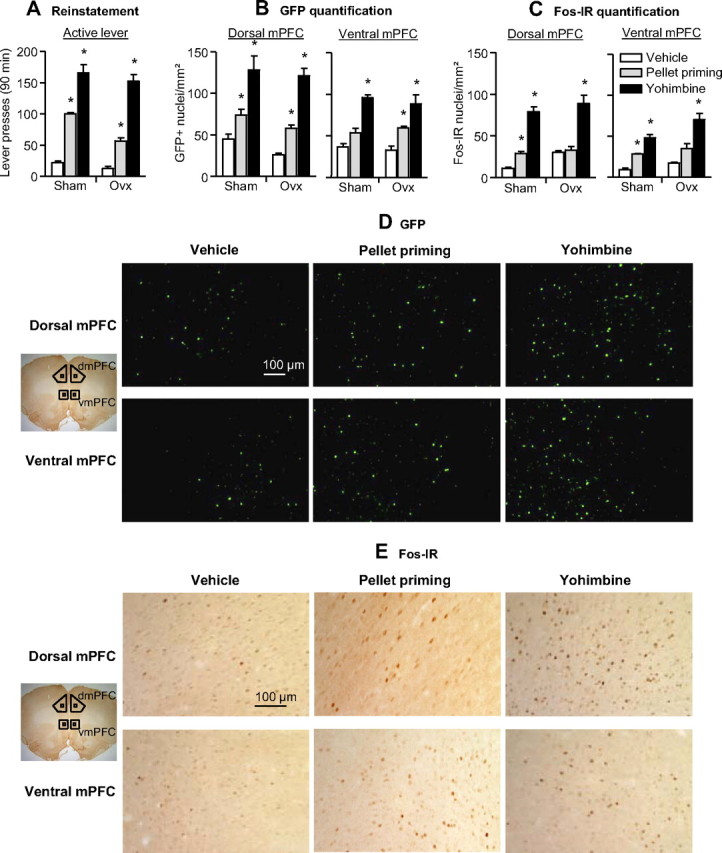
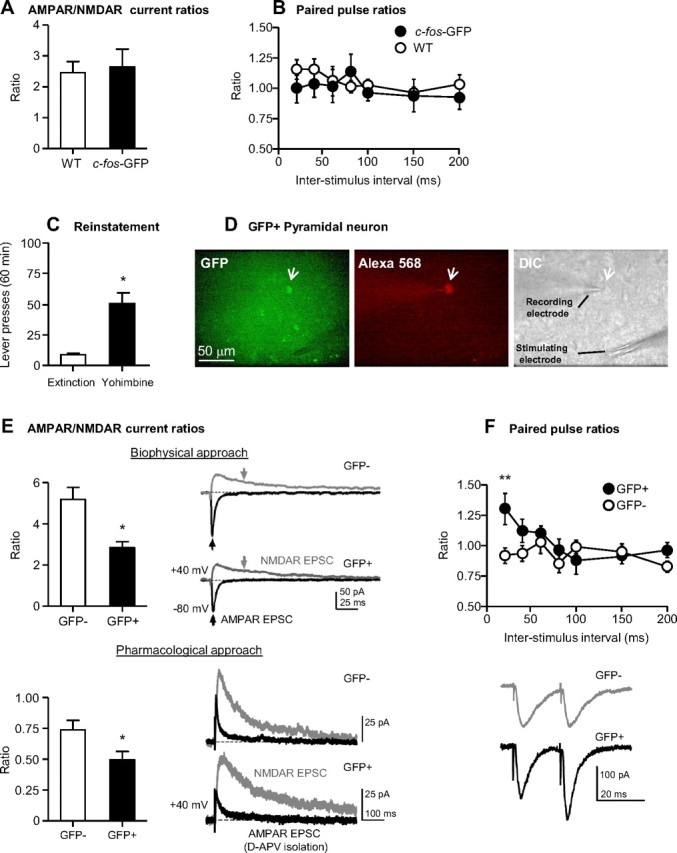
Relapse to maladaptive eating habits during dieting is often provoked by stress and there is evidence for a role of ovarian hormones in stress responses and feeding. We studied the role of these hormones in stress-induced reinstatement of food seeking and medial prefrontal cortex (mPFC) neuronal activation in c-fos-GFP transgenic female rats, which express GFP in strongly activated neurons. Food-restricted ovariectomized or sham-operated c-fos-GFP rats were trained to lever-press for palatable food pellets. Subsequently, lever-pressing was extinguished and reinstatement of food seeking and mPFC neuronal activation was assessed after injections of the pharmacological stressor yohimbine (0.5-2 mg/kg) or pellet priming (1-4 noncontingent pellets). Estrous cycle effects on reinstatement were also assessed in wild-type rats. Yohimbine- and pellet-priming-induced reinstatement was associated with Fos and GFP induction in mPFC; both reinstatement and neuronal activation were minimally affected by ovarian hormones in both c-fos-GFP and wild-type rats. c-fos-GFP transgenic rats were then used to assess glutamatergic synaptic alterations within activated GFP-positive and nonactivated GFP-negative mPFC neurons following yohimbine-induced reinstatement of food seeking. This reinstatement was associated with reduced AMPA receptor/NMDA receptor current ratios and increased paired-pulse facilitation in activated GFP-positive but not GFP-negative neurons. While ovarian hormones do not appear to play a role in stress-induced relapse of food seeking in our rat model, this reinstatement was associated with unique synaptic alterations in strongly activated mPFC neurons. Our paper introduces the c-fos-GFP transgenic rat as a new tool to study unique synaptic changes in activated neurons during behavior.
Figures
References
-
- Anzalone CR, Hong LS, Lu JK, LaPolt PS. Influences of age and ovarian follicular reserve on estrous cycle patterns, ovulation, and hormone secretion in the Long-Evans rat. Biol Reprod. 2001;64:1056–1062. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical