

Activity-dependent growth of new dendritic spines is regulated by the proteasome
- PMID: 22726833
- PMCID: PMC3500563
- DOI: 10.1016/j.neuron.2012.04.031
Activity-dependent growth of new dendritic spines is regulated by the proteasome
Abstract
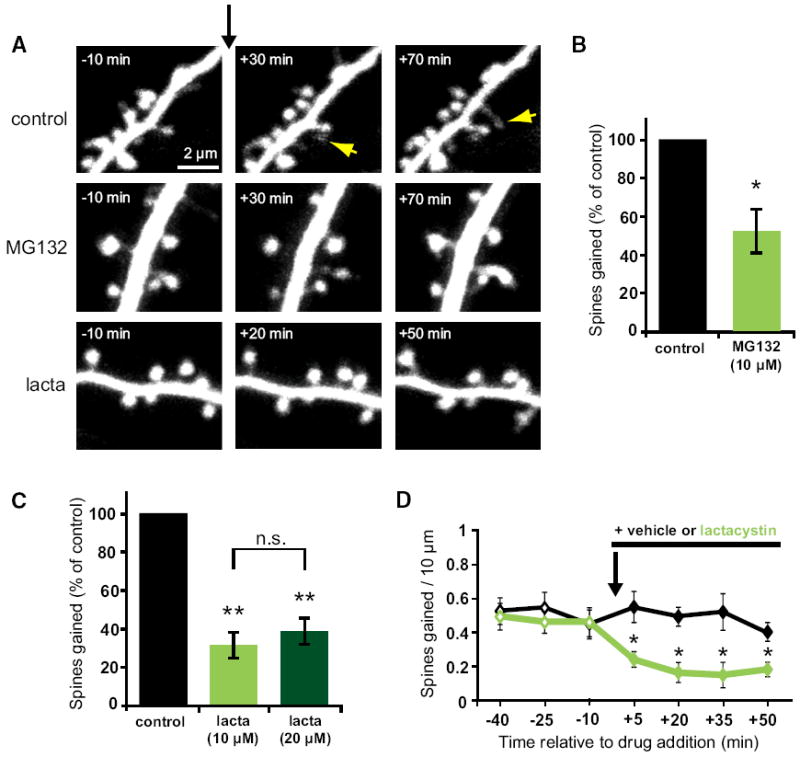
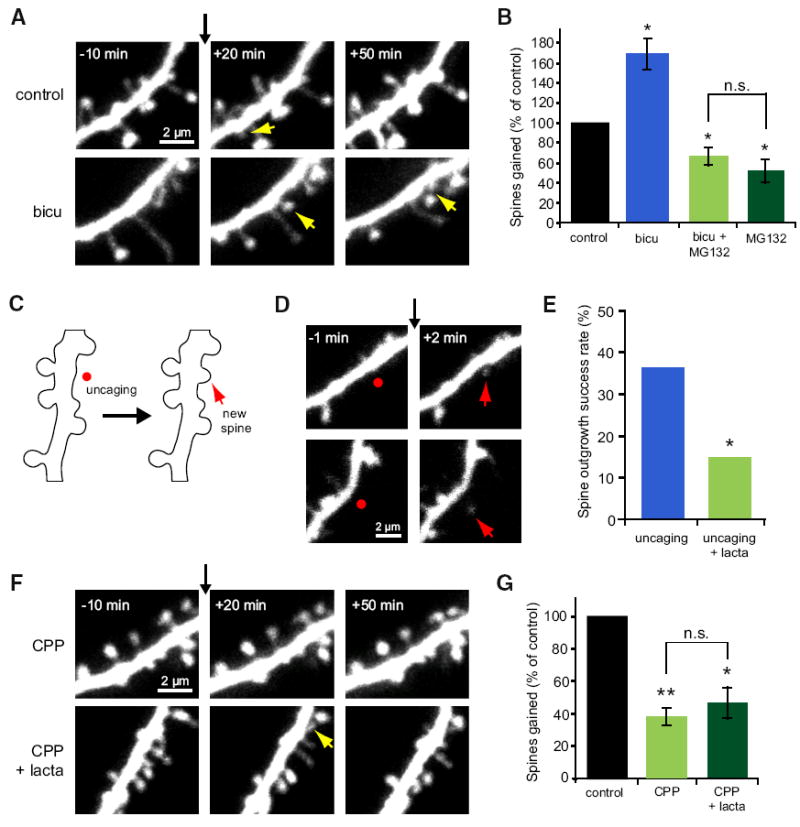
Growth of new dendritic spines contributes to experience-dependent circuit plasticity in the cerebral cortex. Yet the signaling mechanisms leading to new spine outgrowth remain poorly defined. Increasing evidence supports that the proteasome is an important mediator of activity-dependent neuronal signaling. We therefore tested the role of the proteasome in activity-dependent spinogenesis. Using pharmacological manipulations, glutamate uncaging, and two-photon imaging of GFP-transfected hippocampal pyramidal neurons, we demonstrate that acute inhibition of the proteasome blocks activity-induced spine outgrowth. Remarkably, mutation of serine 120 to alanine of the Rpt6 proteasomal subunit in individual neurons was sufficient to block activity-induced spine outgrowth. Signaling through NMDA receptors and CaMKII, but not PKA, is required to facilitate spine outgrowth. Moreover, abrogating CaMKII binding to the NMDA receptor abolished activity-induced spinogenesis. Our data support a model in which neural activity facilitates spine outgrowth via an NMDA receptor- and CaMKII-dependent increase in local proteasomal degradation.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures


References
-
- Acconcia F, Sigismund S, Polo S. Ubiquitin in trafficking: the network at work. Exp Cell Res. 2009;315:1610–1618. - PubMed
-
- Bhatt DH, Zhang S, Gan WB. Dendritic spine dynamics. Annu Rev Physiol. 2009;71:261–282. - PubMed
-
- Bingol B, Schuman EM. Activity-dependent dynamics and sequestration of proteasomes in dendritic spines. Nature. 2006;441:1144–1148. - PubMed
-
- Bingol B, Sheng M. Deconstruction for reconstruction: the role of proteolysis in neural plasticity and disease. Neuron. 2011;69:22–32. - PubMed
-
- Bingol B, Wang CF, Arnott D, Cheng D, Peng J, Sheng M. Autophosphorylated CaMKIIalpha acts as a scaffold to recruit proteasomes to dendritic spines. Cell. 2010;140:567–578. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R25 GM056765/GM/NIGMS NIH HHS/United States
- T32GM007377/GM/NIGMS NIH HHS/United States
- NS054732/NS/NINDS NIH HHS/United States
- AG017502/AG/NIA NIH HHS/United States
- GM056765/GM/NIGMS NIH HHS/United States
- R01 NS062736/NS/NINDS NIH HHS/United States
- R01 AG017502/AG/NIA NIH HHS/United States
- 52005892/HHMI/Howard Hughes Medical Institute/United States
- T34 GM083894/GM/NIGMS NIH HHS/United States
- NS062736/NS/NINDS NIH HHS/United States
- R01 NS060847/NS/NINDS NIH HHS/United States
- R56 AG017502/AG/NIA NIH HHS/United States
- GM083894/GM/NIGMS NIH HHS/United States
- T32 GM007377/GM/NIGMS NIH HHS/United States
- R21 NS054732/NS/NINDS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
