

Oxidative stress fuels Trypanosoma cruzi infection in mice
- PMID: 22728935
- PMCID: PMC3386808
- DOI: 10.1172/JCI58525
Oxidative stress fuels Trypanosoma cruzi infection in mice
Abstract
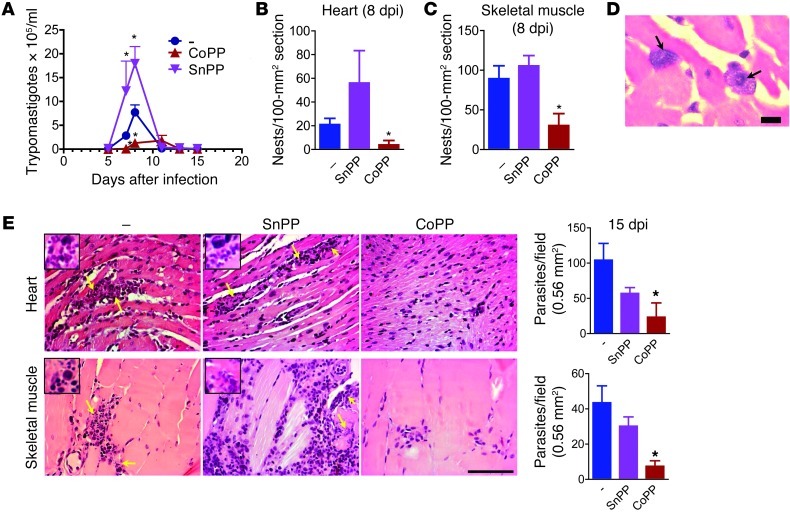
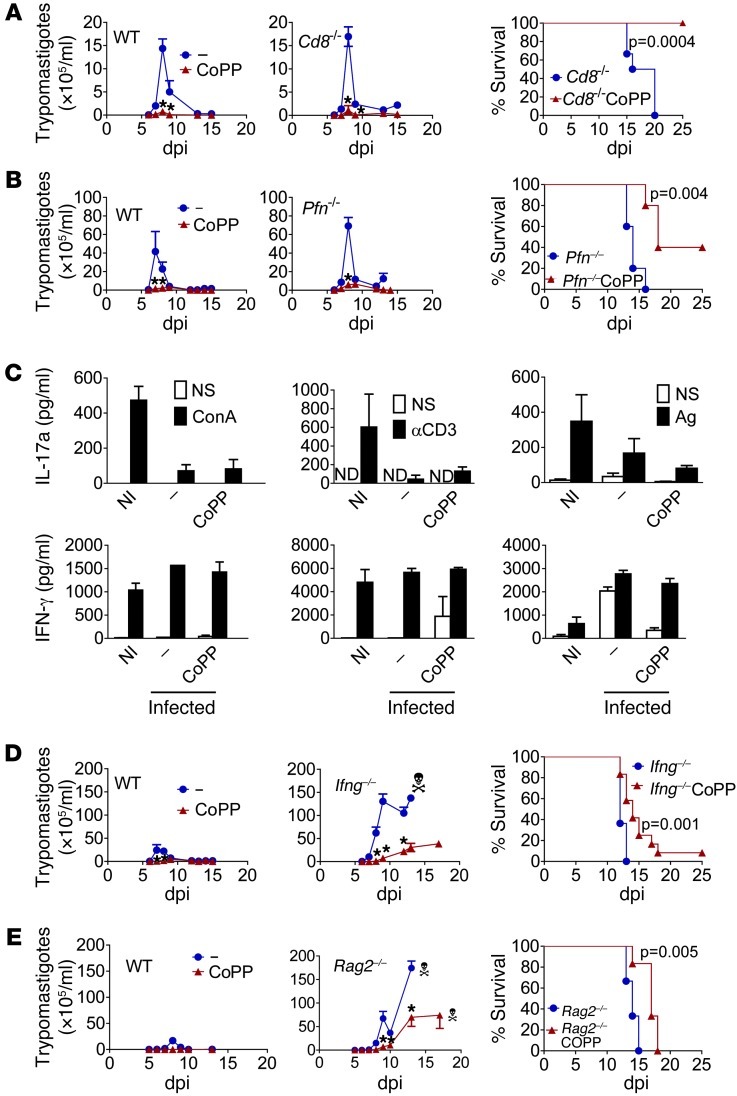
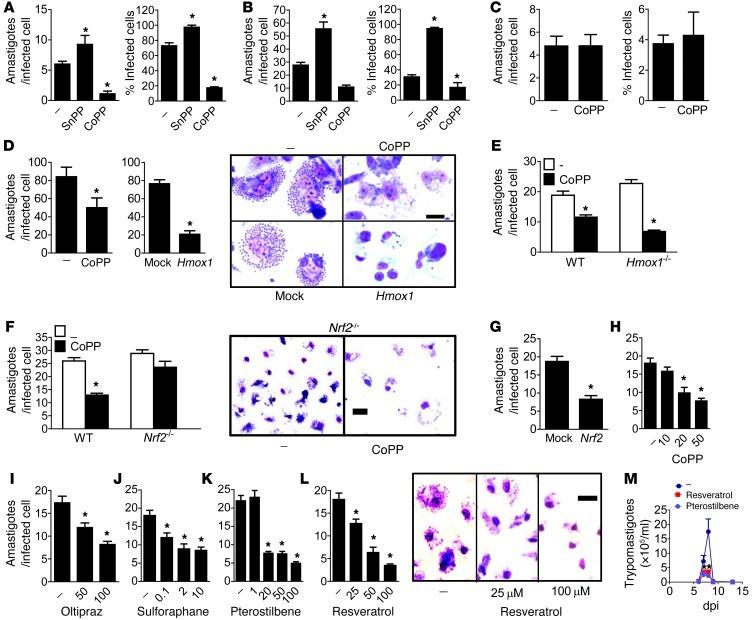
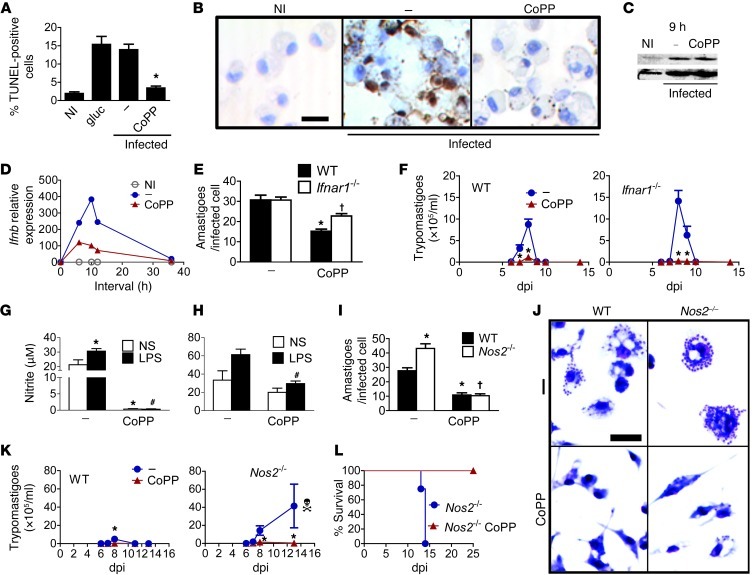
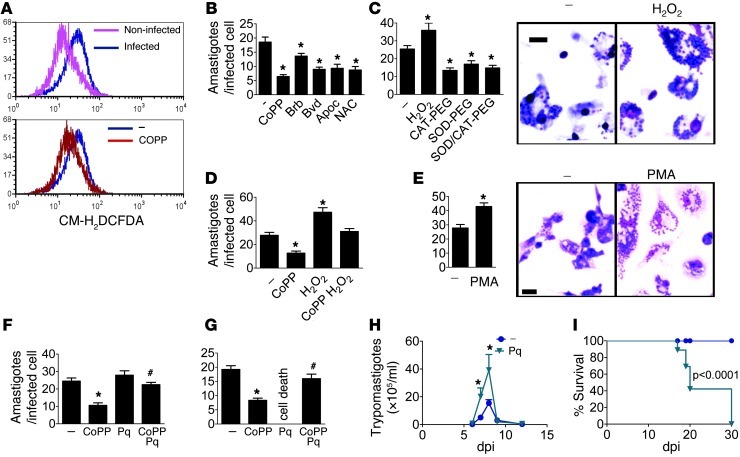
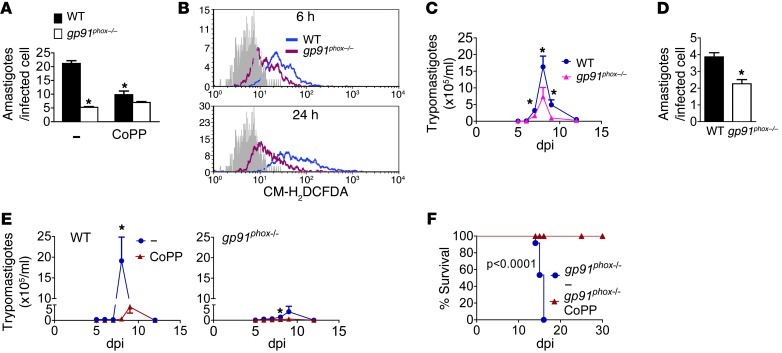
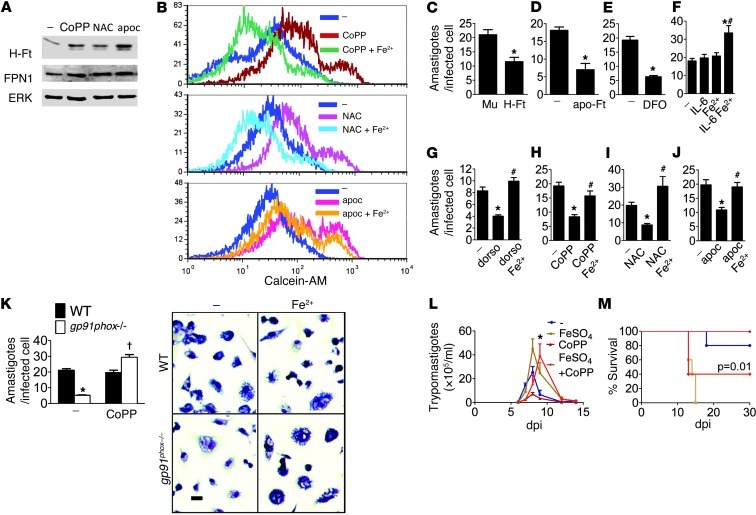
Oxidative damage contributes to microbe elimination during macrophage respiratory burst. Nuclear factor, erythroid-derived 2, like 2 (NRF2) orchestrates antioxidant defenses, including the expression of heme-oxygenase-1 (HO-1). Unexpectedly, the activation of NRF2 and HO-1 reduces infection by a number of pathogens, although the mechanism responsible for this effect is largely unknown. We studied Trypanosoma cruzi infection in mice in which NRF2/HO-1 was induced with cobalt protoporphyrin (CoPP). CoPP reduced parasitemia and tissue parasitism, while an inhibitor of HO-1 activity increased T. cruzi parasitemia in blood. CoPP-induced effects did not depend on the adaptive immunity, nor were parasites directly targeted. We also found that CoPP reduced macrophage parasitism, which depended on NRF2 expression but not on classical mechanisms such as apoptosis of infected cells, induction of type I IFN, or NO. We found that exogenous expression of NRF2 or HO-1 also reduced macrophage parasitism. Several antioxidants, including NRF2 activators, reduced macrophage parasite burden, while pro-oxidants promoted it. Reducing the intracellular labile iron pool decreased parasitism, and antioxidants increased the expression of ferritin and ferroportin in infected macrophages. Ferrous sulfate reversed the CoPP-induced decrease in macrophage parasite burden and, given in vivo, reversed their protective effects. Our results indicate that oxidative stress contributes to parasite persistence in host tissues and open a new avenue for the development of anti-T. cruzi drugs.
Figures
Comment in
-
Oxidative stress and intracellular infections: more iron to the fire.J Clin Invest. 2012 Jul;122(7):2352-4. doi: 10.1172/JCI64239. Epub 2012 Jun 25. J Clin Invest. 2012. PMID: 22728929 Free PMC article.
References
-
- McCord JM. Iron, free radicals, and oxidative injury. J Nutr. 2004;134(11):3171S–3172S. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
