

Acclimatory responses of Arabidopsis to fluctuating light environment: comparison of different sunfleck regimes and accessions
- PMID: 22729524
- PMCID: PMC3430843
- DOI: 10.1007/s11120-012-9757-2
Acclimatory responses of Arabidopsis to fluctuating light environment: comparison of different sunfleck regimes and accessions
Abstract
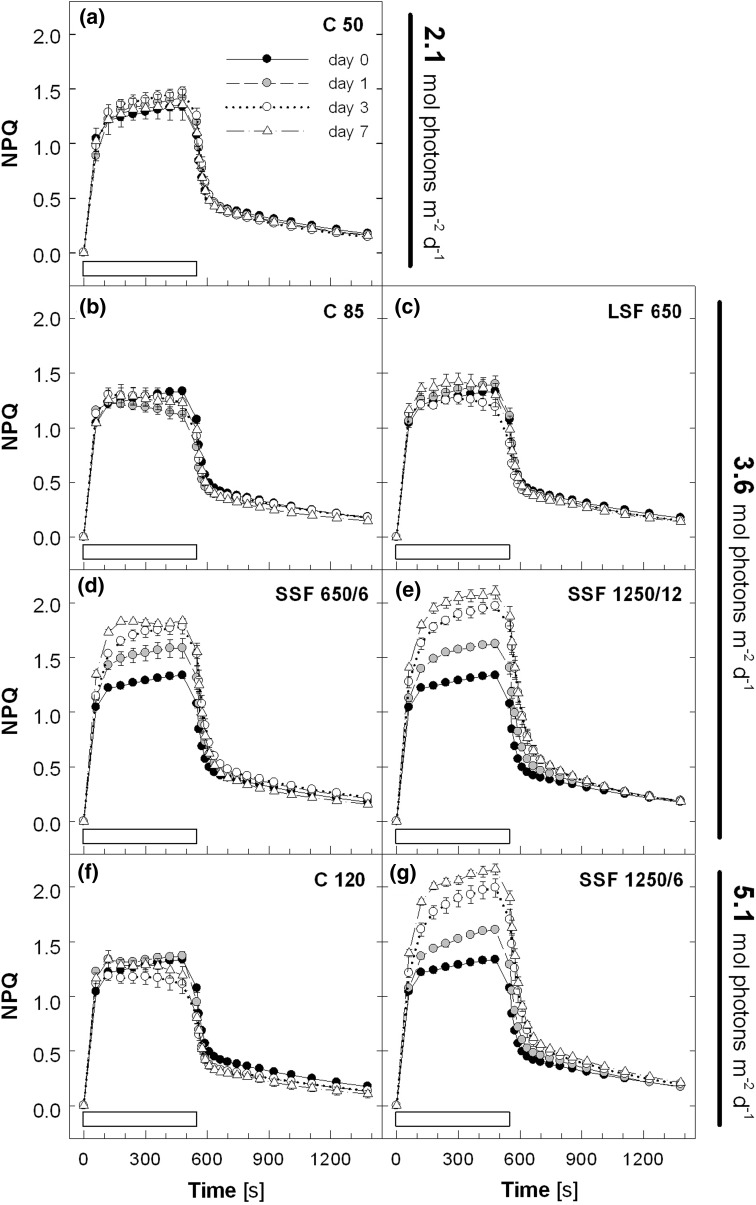
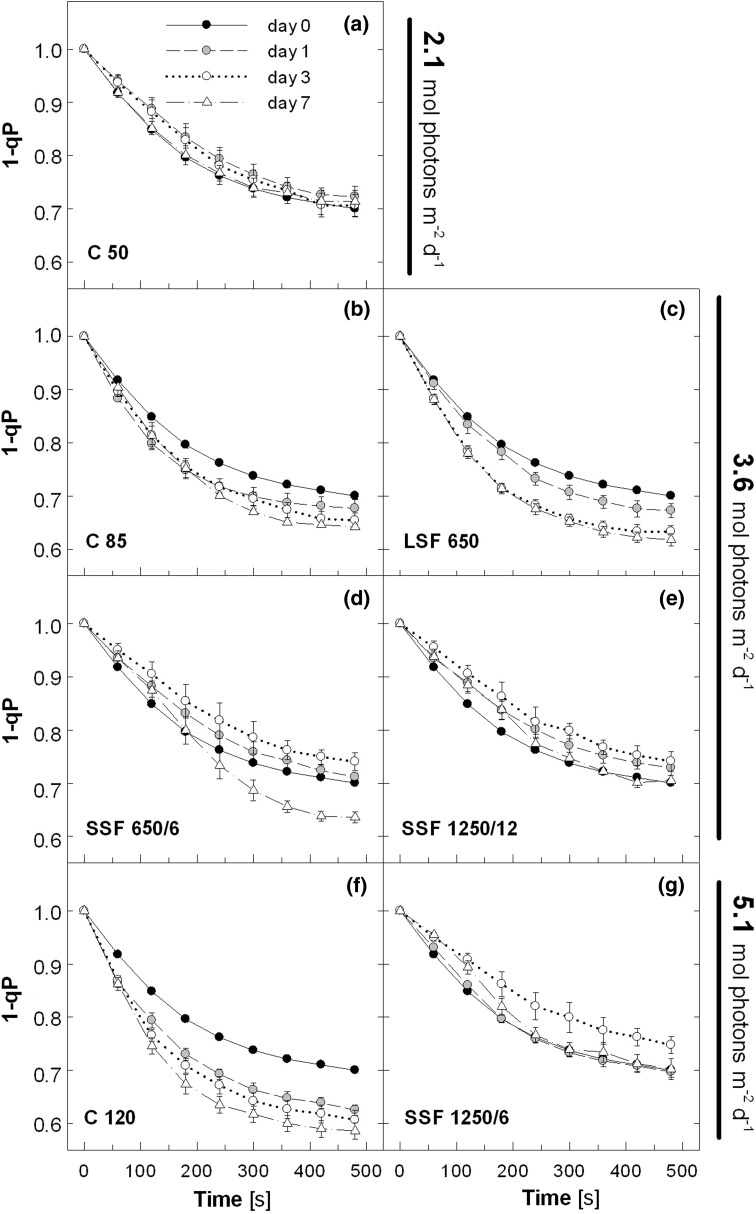
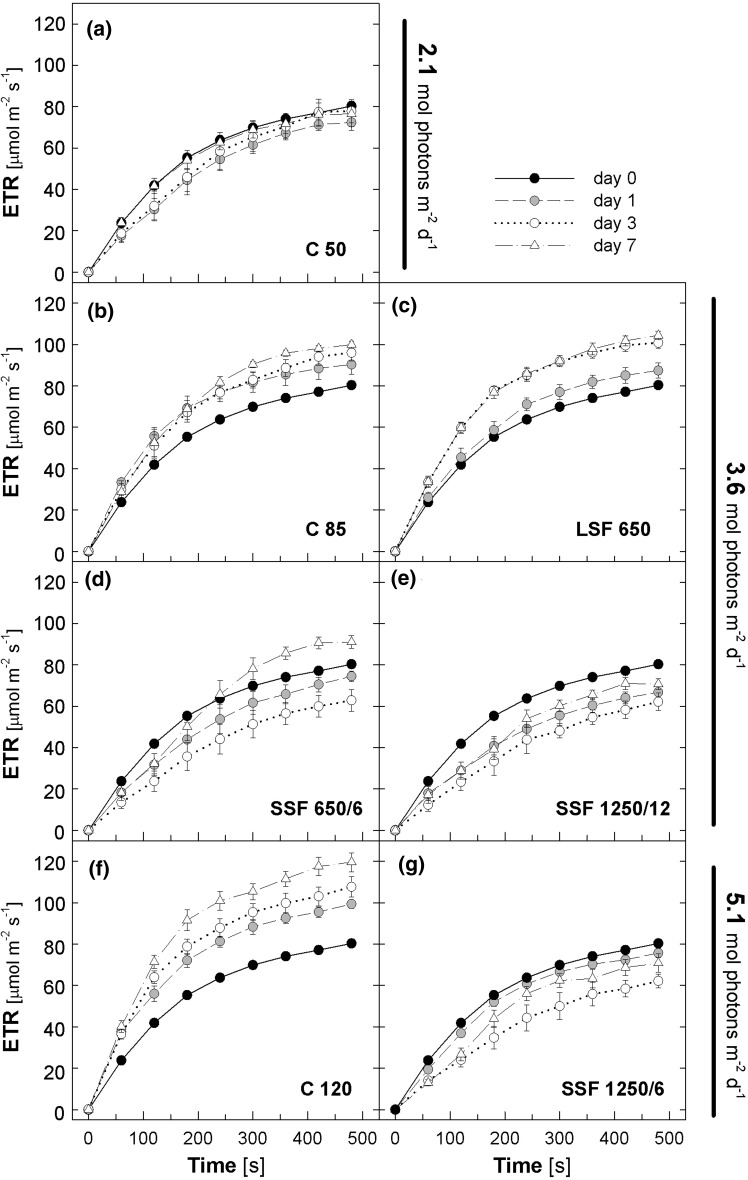
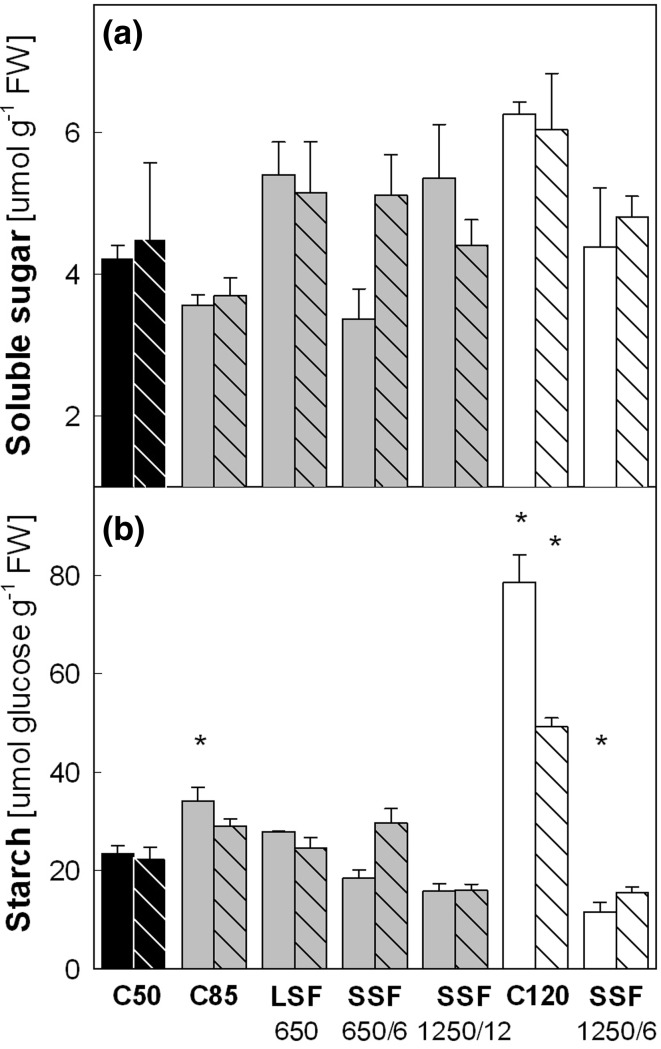
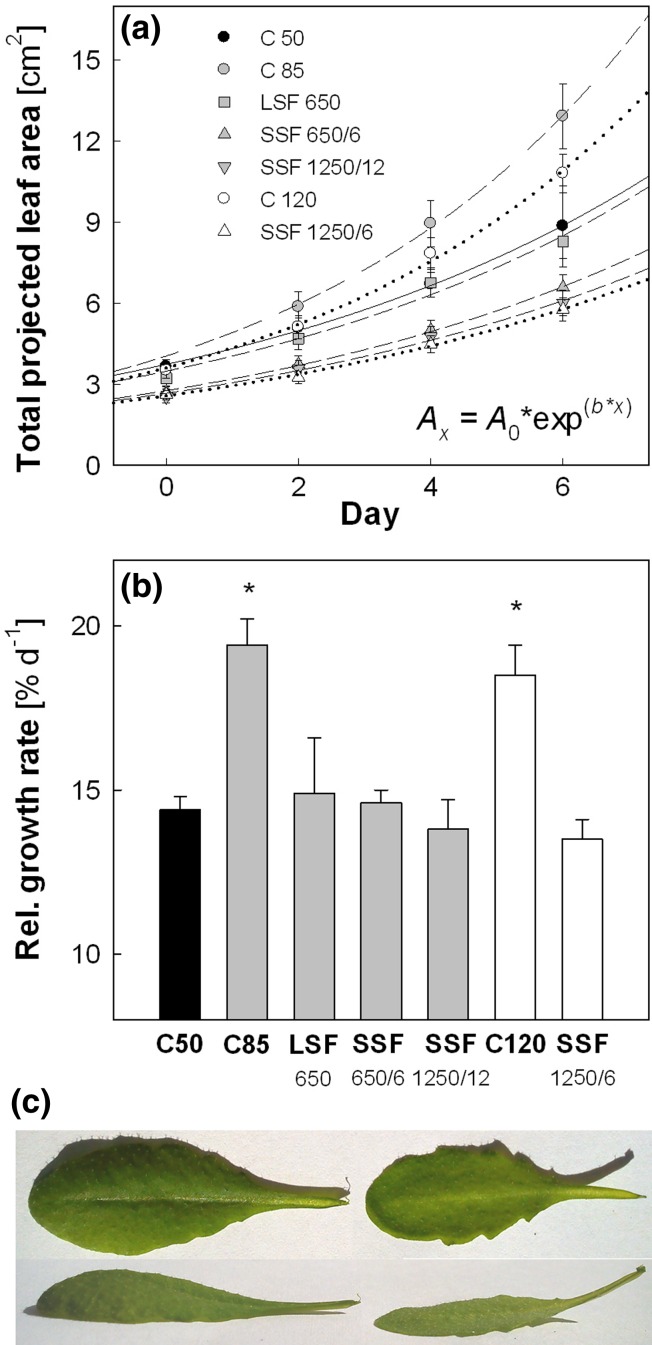
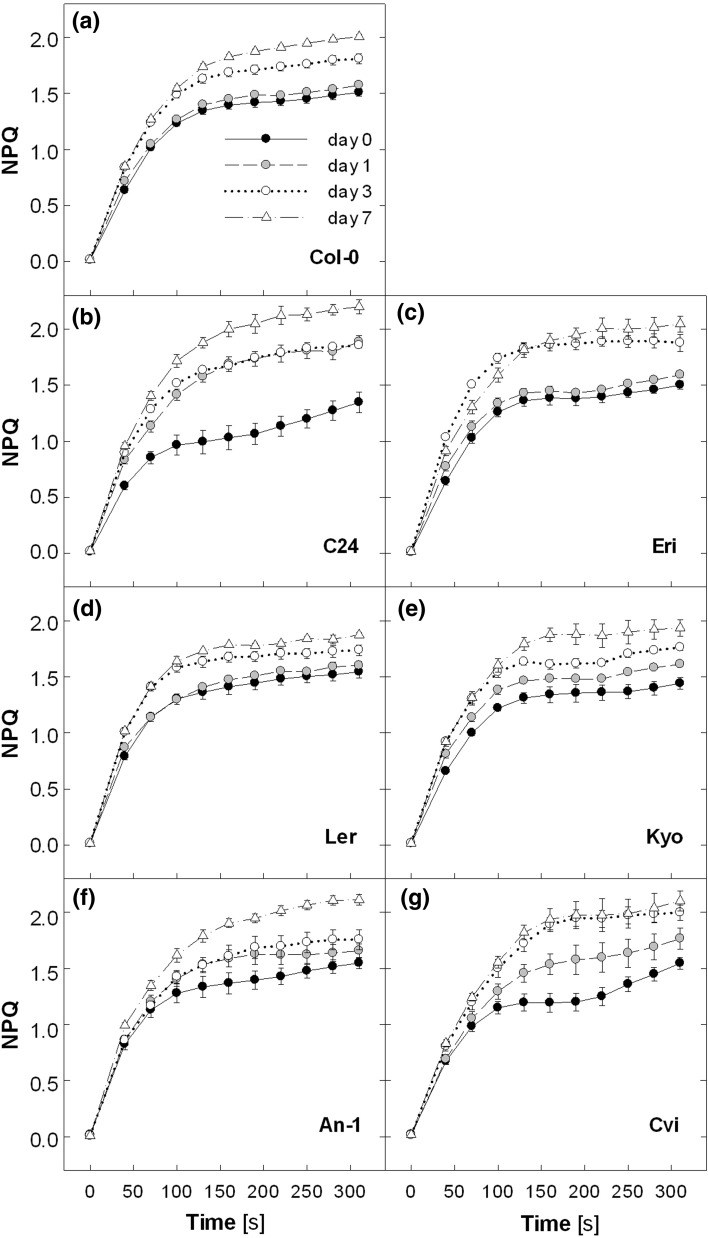
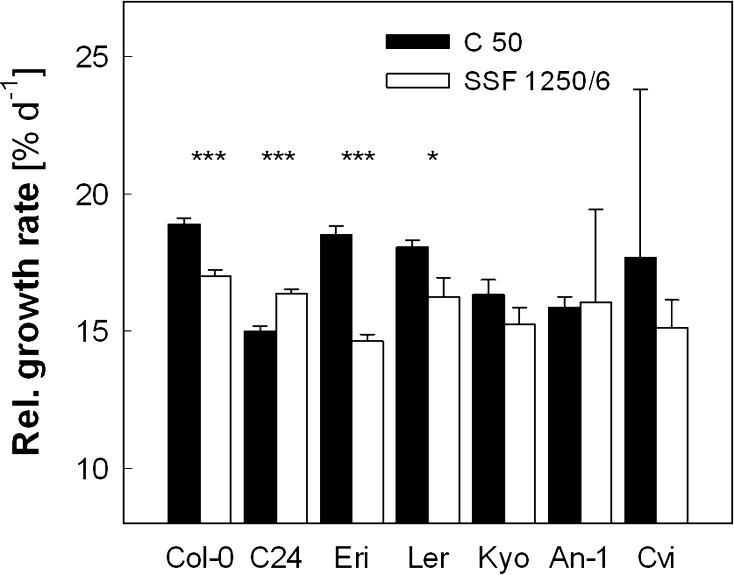
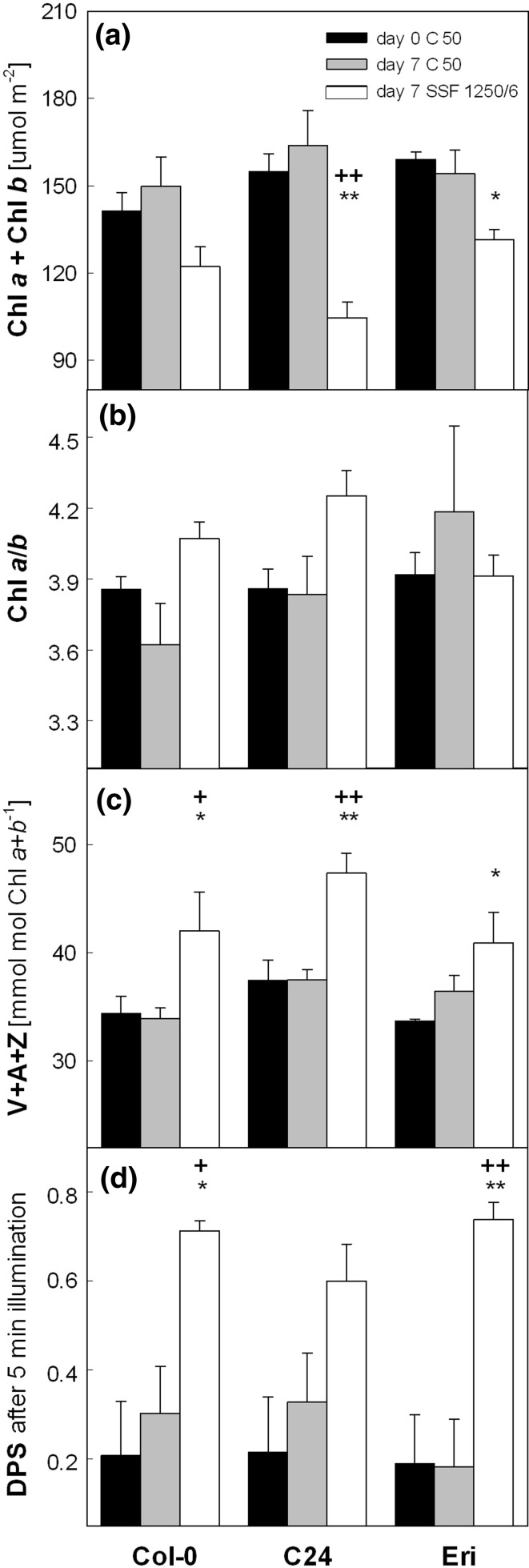
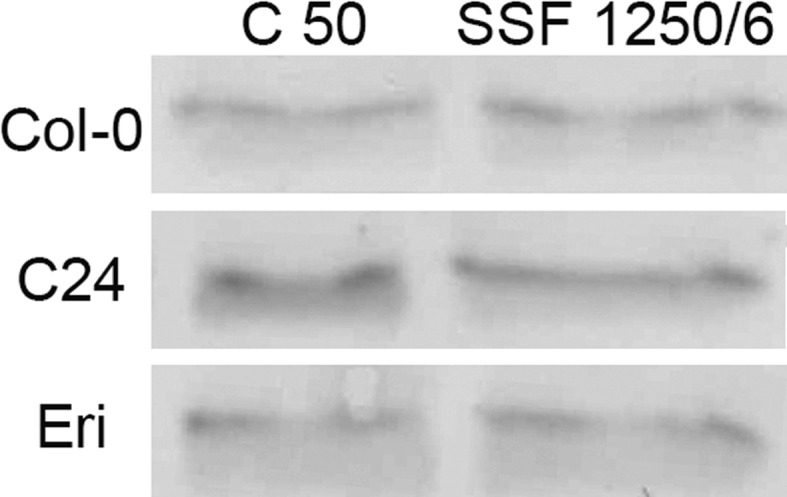
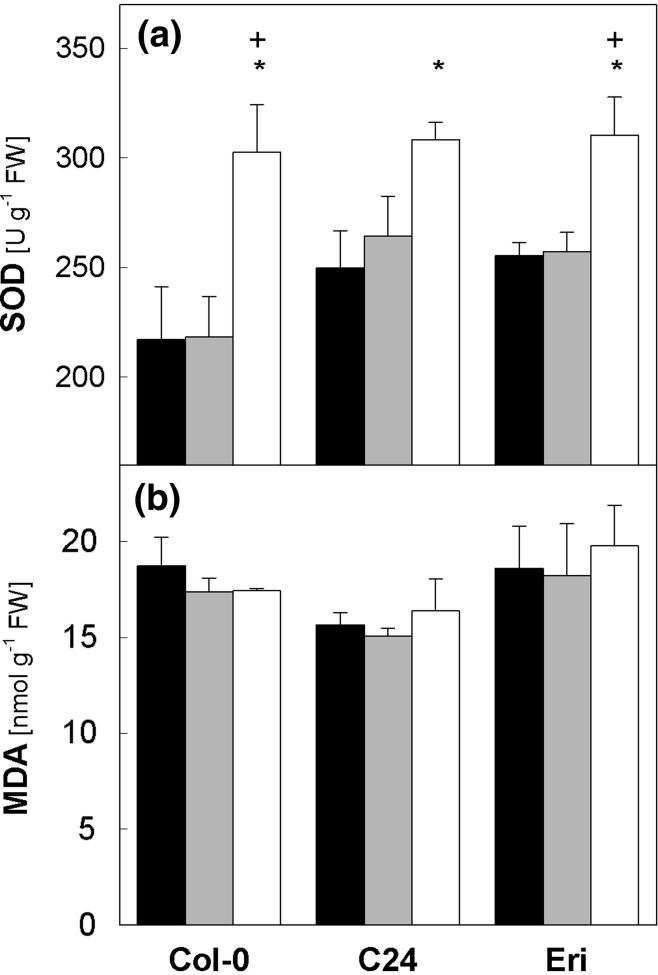
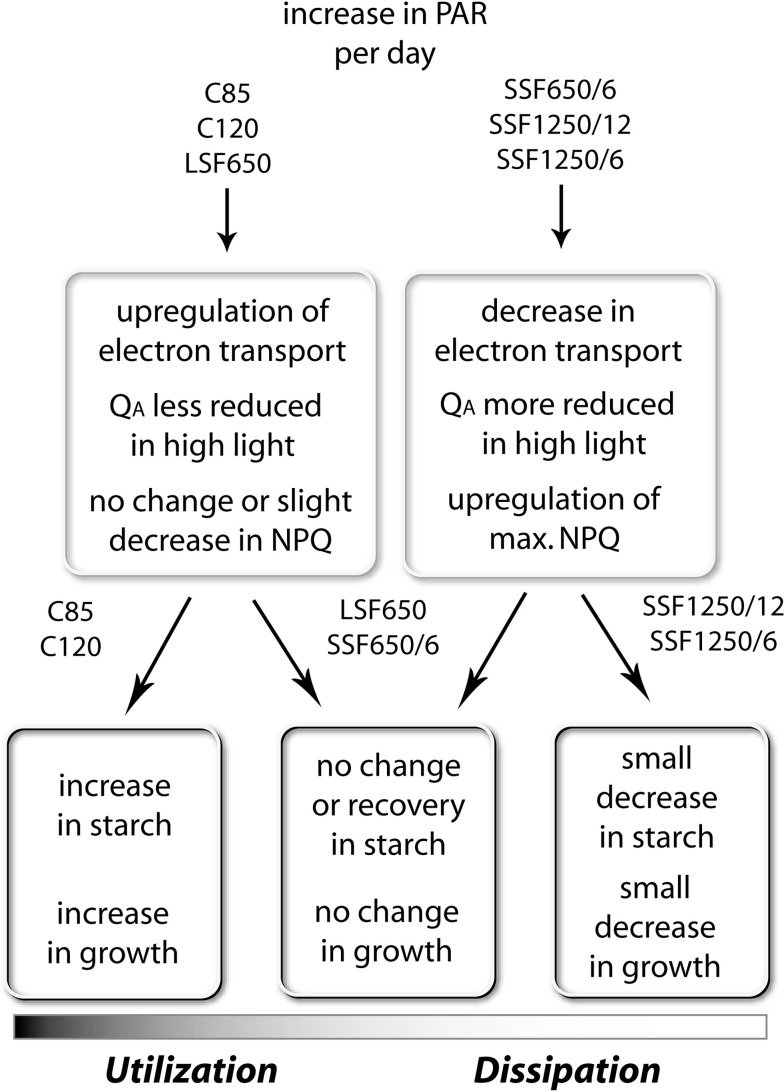
Acclimation to fluctuating light environment with short (lasting 20 s, at 650 or 1,250 μmol photons m(-2) s(-1), every 6 or 12 min) or long (for 40 min at 650 μmol photons m(-2) s(-1), once a day at midday) sunflecks was studied in Arabidopsis thaliana. The sunfleck treatments were applied in the background daytime light intensity of 50 μmol photons m(-2) s(-1). In order to distinguish the effects of sunflecks from those of increased daily irradiance, constant light treatments at 85 and 120 μmol photons m(-2) s(-1), which gave the same photosynthetically active radiation (PAR) per day as the different sunfleck treatments, were also included in the experiments. The increased daily total PAR in the two higher constant light treatments enhanced photosystem II electron transport and starch accumulation in mature leaves and promoted expansion of young leaves in Columbia-0 plants during the 7-day treatments. Compared to the plants remaining under 50 μmol photons m(-2) s(-1), application of long sunflecks caused upregulation of electron transport without affecting carbon gain in the form of starch accumulation and leaf growth or the capacity of non-photochemical quenching (NPQ). Mature leaves showed marked enhancement of the NPQ capacity under the conditions with short sunflecks, which preceded recovery and upregulation of electron transport, demonstrating the initial priority of photoprotection. The distinct acclimatory responses to constant PAR, long sunflecks, and different combinations of short sunflecks are consistent with acclimatory adjustment of the processes in photoprotection and carbon gain, depending on the duration, frequency, and intensity of light fluctuations. While the responses of leaf expansion to short sunflecks differed among the seven Arabidopsis accessions examined, all plants showed NPQ upregulation, suggesting limited ability of this species to utilize short sunflecks. The increase in the NPQ capacity was accompanied by reduced chlorophyll contents, higher levels of the xanthophyll-cycle pigments, faster light-induced de-epoxidation of violaxanthin to zeaxanthin and antheraxanthin, increased amounts of PsbS protein, as well as enhanced activity of superoxide dismutase. These acclimatory mechanisms, involving reorganization of pigment-protein complexes and upregulation of other photoprotective reactions, are probably essential for Arabidopsis plants to cope with photo-oxidative stress induced by short sunflecks without suffering from severe photoinhibition and lipid peroxidation.
Figures
References
-
- Adams WWIII, Demmig-Adams B, Logan BA, Barker DH, Osmond CB. Rapid changes in xanthophyll cycle-dependent energy dissipation and photosystem II efficiency in two vines, Stephania japonica and Smilax australis, growing in the understory of an open Eucalyptus forest. Plant Cell Environ. 1999;22:125–136. doi: 10.1046/j.1365-3040.1999.00369.x. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
