

Fluorescence analysis of the lipid binding-induced conformational change of apolipoprotein E4
- PMID: 22730894
- PMCID: PMC3447985
- DOI: 10.1021/bi300672s
Fluorescence analysis of the lipid binding-induced conformational change of apolipoprotein E4
Abstract
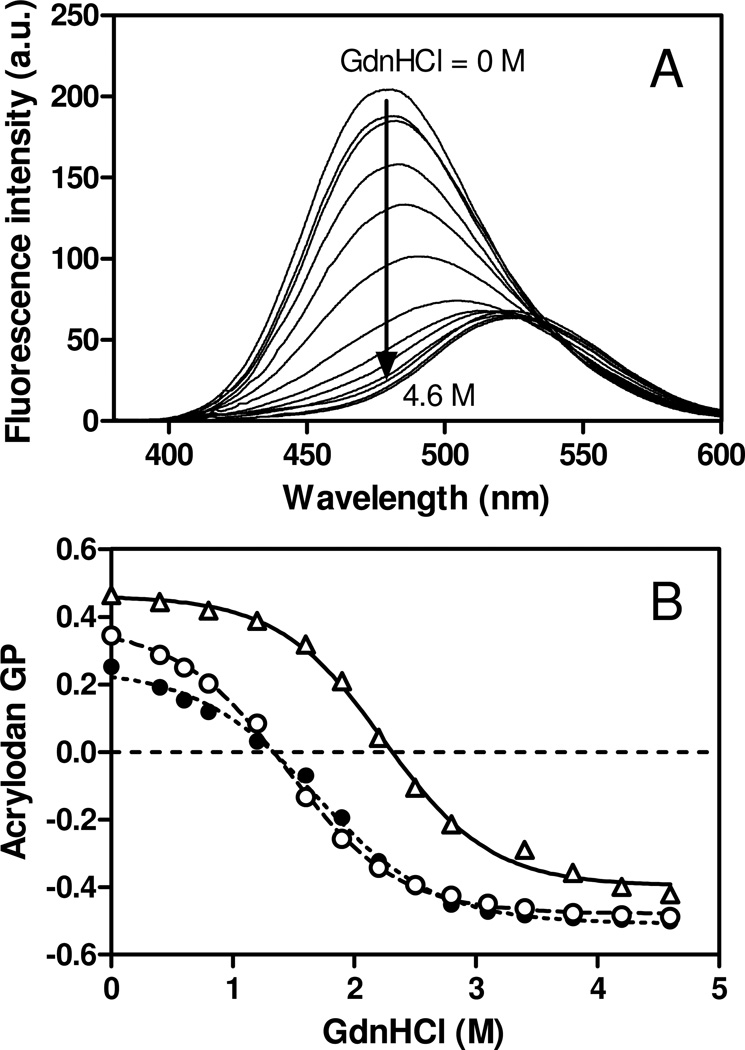
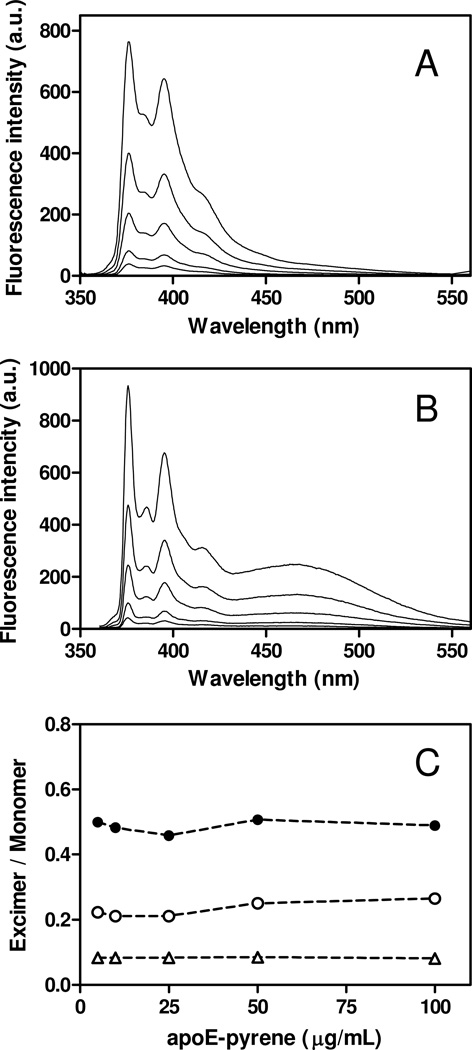
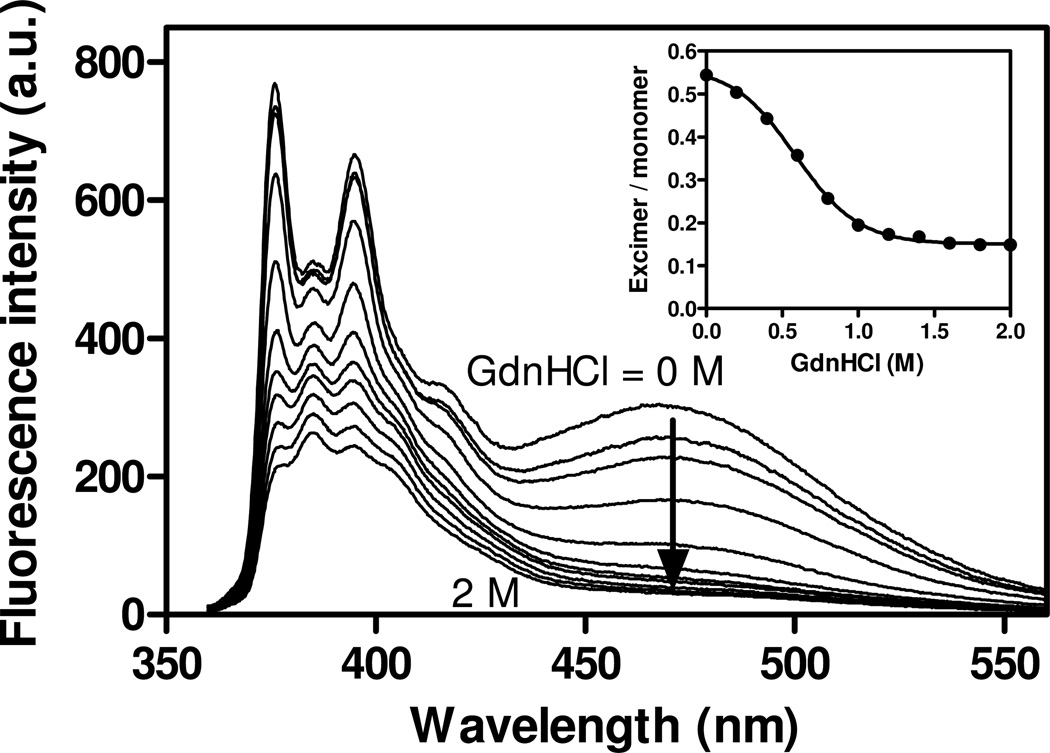
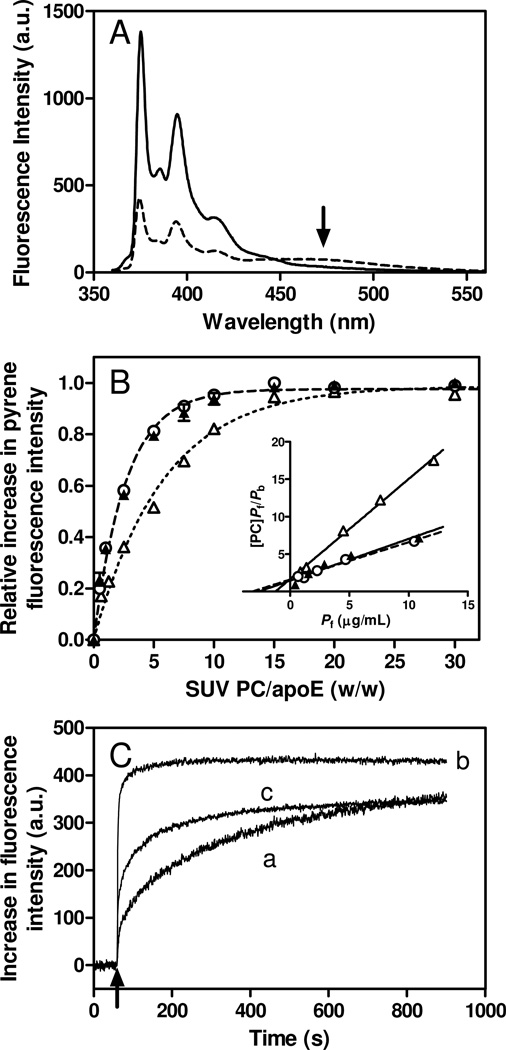
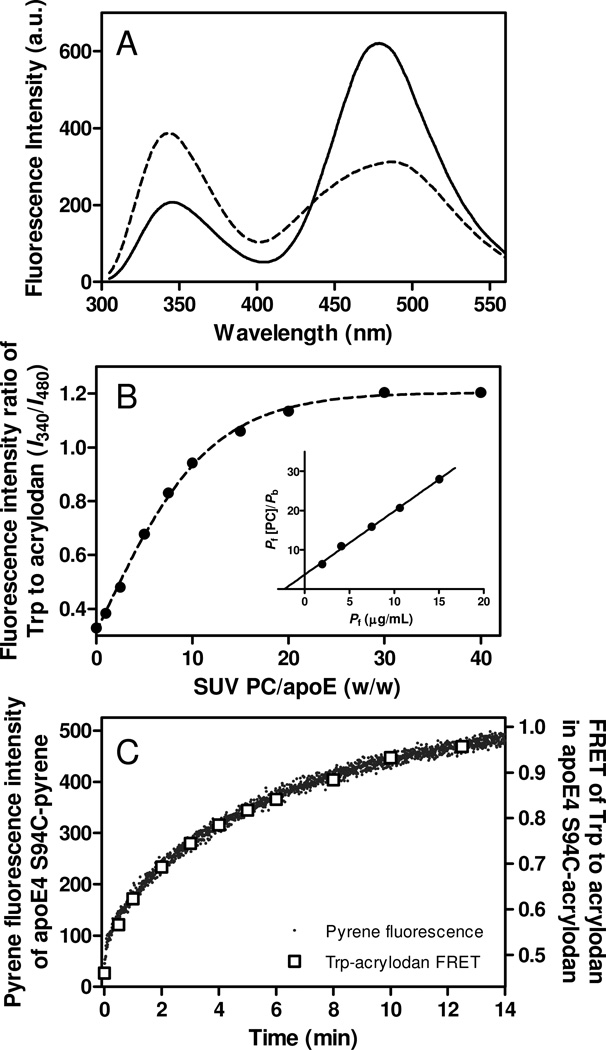
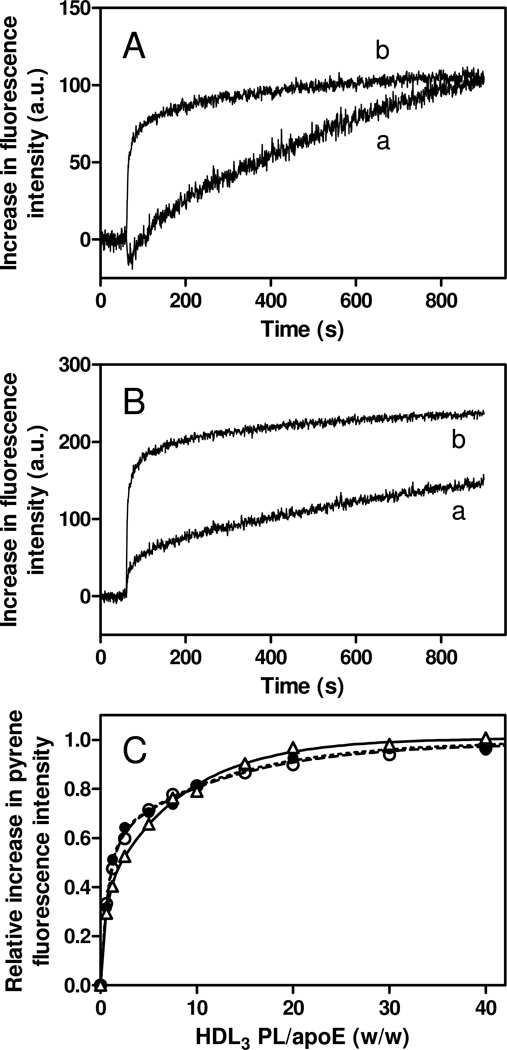
Apolipoprotein (apo) E is thought to undergo conformational changes in the N-terminal helix bundle domain upon lipid binding, modulating its receptor binding activity. In this study, site-specific fluorescence labeling of the N-terminal (S94) and C-terminal (W264 or S290) helices in apoE4 by pyrene maleimide or acrylodan was employed to probe the conformational organization and lipid binding behavior of the N- and C-terminal domains. Guanidine denaturation experiments monitored by acrylodan fluorescence demonstrated the less organized, more solvent-exposed structure of the C-terminal helices compared to the N-terminal helix bundle. Pyrene excimer fluorescence together with gel filtration chromatography indicated that there are extensive intermolecular helix-helix contacts through the C-terminal helices of apoE4. Comparison of increases in pyrene fluorescence upon binding of pyrene-labeled apoE4 to egg phosphatidylcholine small unilamellar vesicles suggests a two-step lipid-binding process; apoE4 initially binds to a lipid surface through the C-terminal helices followed by the slower conformational reorganization of the N-terminal helix bundle domain. Consistent with this, fluorescence resonance energy transfer measurements from Trp residues to acrylodan attached at position 94 demonstrated that upon binding to the lipid surface, opening of the N-terminal helix bundle occurs at the same rate as the increase in pyrene fluorescence of the N-terminal domain. Such a two-step mechanism of lipid binding of apoE4 is likely to apply to mostly phospholipid-covered lipoproteins such as VLDL. However, monitoring pyrene fluorescence upon binding to HDL(3) suggests that not only apoE-lipid interactions but also protein-protein interactions are important for apoE4 binding to HDL(3).
Figures
Similar articles
-
Fluorescence study of domain structure and lipid interaction of human apolipoproteins E3 and E4.Biochim Biophys Acta. 2014 Dec;1841(12):1716-24. doi: 10.1016/j.bbalip.2014.09.019. Biochim Biophys Acta. 2014. PMID: 25281910 Free PMC article.
-
Contributions of the carboxyl-terminal helical segment to the self-association and lipoprotein preferences of human apolipoprotein E3 and E4 isoforms.Biochemistry. 2008 Mar 4;47(9):2968-77. doi: 10.1021/bi701923h. Epub 2008 Jan 18. Biochemistry. 2008. PMID: 18201068 Free PMC article.
-
Examination of lipid-bound conformation of apolipoprotein E4 by pyrene excimer fluorescence.J Biol Chem. 2005 Apr 15;280(15):14605-10. doi: 10.1074/jbc.M414019200. Epub 2005 Feb 11. J Biol Chem. 2005. PMID: 15708851
-
Apolipoprotein E isoforms and lipoprotein metabolism.IUBMB Life. 2014 Sep;66(9):616-23. doi: 10.1002/iub.1314. IUBMB Life. 2014. PMID: 25328986 Review.
-
Apolipoprotein A-I: structure-function relationships.J Lipid Res. 2000 Jun;41(6):853-72. J Lipid Res. 2000. PMID: 10828078 Review.
Cited by
-
Molecular mechanisms responsible for the differential effects of apoE3 and apoE4 on plasma lipoprotein-cholesterol levels.Arterioscler Thromb Vasc Biol. 2013 Apr;33(4):687-93. doi: 10.1161/ATVBAHA.112.301193. Epub 2013 Feb 14. Arterioscler Thromb Vasc Biol. 2013. PMID: 23413428 Free PMC article.
-
The LDL receptor binding domain of apolipoprotein E directs the relative orientation of its C-terminal segment in reconstituted nascent HDL.Biochim Biophys Acta Biomembr. 2021 Jul 1;1863(7):183618. doi: 10.1016/j.bbamem.2021.183618. Epub 2021 Apr 6. Biochim Biophys Acta Biomembr. 2021. PMID: 33831404 Free PMC article.
-
ApoE: In Vitro Studies of a Small Molecule Effector.Biochemistry. 2016 May 10;55(18):2613-21. doi: 10.1021/acs.biochem.6b00324. Epub 2016 Apr 27. Biochemistry. 2016. PMID: 27065061 Free PMC article.
-
Fluorescence study of domain structure and lipid interaction of human apolipoproteins E3 and E4.Biochim Biophys Acta. 2014 Dec;1841(12):1716-24. doi: 10.1016/j.bbalip.2014.09.019. Biochim Biophys Acta. 2014. PMID: 25281910 Free PMC article.
-
Amyloidogenic Mutation Promotes Fibril Formation of the N-terminal Apolipoprotein A-I on Lipid Membranes.J Biol Chem. 2015 Aug 21;290(34):20947-20959. doi: 10.1074/jbc.M115.664227. Epub 2015 Jul 14. J Biol Chem. 2015. PMID: 26175149 Free PMC article.
References
-
- Mahley RW, Ji ZS. Remnant lipoprotein metabolism: key pathways involving cell-surface heparan sulfate proteoglycans and apolipoprotein E. J. Lipid Res. 1999;40:1–16. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
