

Xenopus laevis zygote arrest 2 (zar2) encodes a zinc finger RNA-binding protein that binds to the translational control sequence in the maternal Wee1 mRNA and regulates translation
- PMID: 22732570
- PMCID: PMC3423596
- DOI: 10.1016/j.ydbio.2012.06.012
Xenopus laevis zygote arrest 2 (zar2) encodes a zinc finger RNA-binding protein that binds to the translational control sequence in the maternal Wee1 mRNA and regulates translation
Abstract
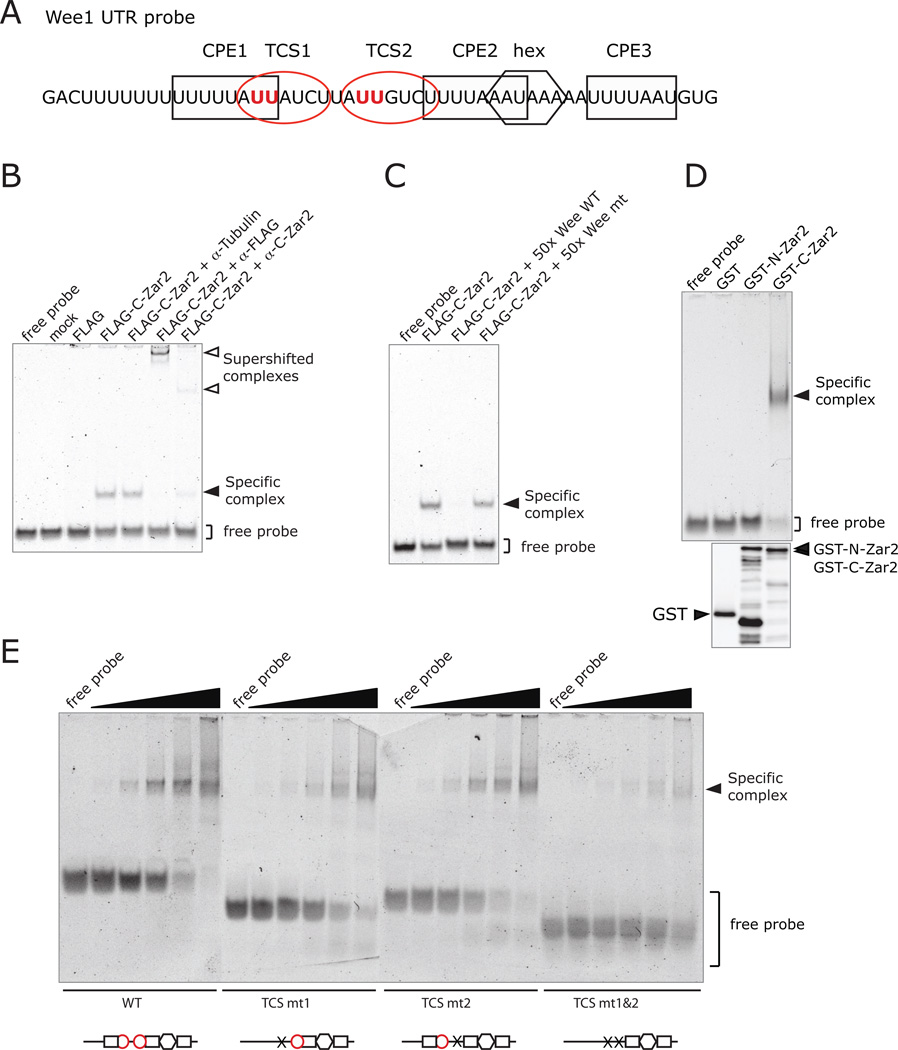
Zygote arrest (Zar) proteins are crucial for early embryonic development, but their molecular mechanism of action is unknown. The Translational Control Sequence (TCS) in the 3' untranslated region (UTR) of the maternal mRNA, Wee1, mediates translational repression in immature Xenopus oocytes and translational activation in mature oocytes, but the protein that binds to the TCS and mediates translational control is not known. Here we show that Xenopus laevis Zar2 (encoded by zar2) binds to the TCS in maternal Wee1 mRNA and represses translation in immature oocytes. Using yeast 3 hybrid assays and electrophoretic mobility shift assays, Zar2 was shown to bind specifically to the TCS in the Wee1 3'UTR. RNA binding required the presence of Zn(2+) and conserved cysteines in the C-terminal domain, suggesting that Zar2 contains a zinc finger. Consistent with regulating maternal mRNAs, Zar2 was present throughout oogenesis, and endogenous Zar2 co-immunoprecipitated endogenous Wee1 mRNA from immature oocytes, demonstrating the physiological significance of the protein-RNA interaction. Interestingly, Zar2 levels decreased during oocyte maturation. Dual luciferase reporter tethered assays showed that Zar2 repressed translation in immature oocytes. Translational repression was relieved during oocyte maturation and this coincided with degradation of Zar2 during maturation. This is the first report of a molecular function of zygote arrest proteins. These data show that Zar2 contains a zinc finger and is a trans-acting factor for the TCS in maternal mRNAs in immature Xenopus oocytes.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures






References
-
- Balzer E, Moss EG. Localization of the developmental timing regulator Lin28 to mRNP complexes, P-bodies and stress granules. RNA Biol. 2007;4:16–25. - PubMed
-
- Bernstein DS, Buter N, Stumpf C, Wickens M. Analyzing mRNA-protein complexes using a yeast three-hybrid system. Methods. 2002;26:123–141. - PubMed
-
- Bienz M. The PHD finger, a nuclear protein-interaction domain. Trends Biochem Sci. 2006;31:35–40. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
