

Research resource: whole transcriptome RNA sequencing detects multiple 1α,25-dihydroxyvitamin D(3)-sensitive metabolic pathways in developing zebrafish
- PMID: 22734042
- PMCID: PMC3434529
- DOI: 10.1210/me.2012-1113
Research resource: whole transcriptome RNA sequencing detects multiple 1α,25-dihydroxyvitamin D(3)-sensitive metabolic pathways in developing zebrafish
Abstract
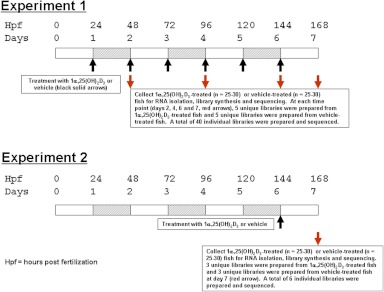
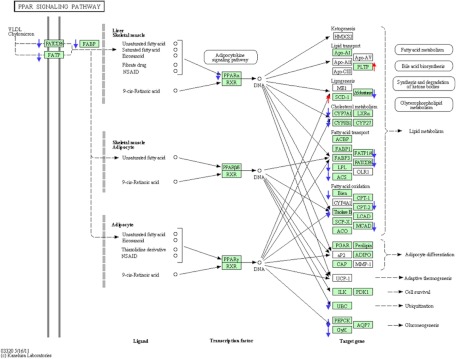
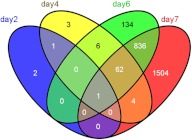
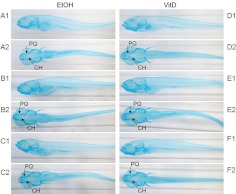
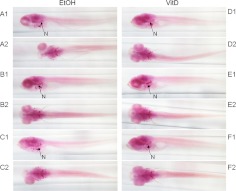
The biological role of vitamin D receptors (VDR), which are abundantly expressed in developing zebrafish (Danio rerio) as early as 48 h after fertilization, and before the development of a mineralized skeleton and mature intestine and kidney, is unknown. We probed the role of VDR in developing zebrafish biology by examining changes in expression of RNA by whole transcriptome shotgun sequencing (RNA-seq) in fish treated with picomolar concentrations of the VDR ligand and hormonal form of vitamin D(3), 1α,25-dihydroxyvitamin D(3) [1α,25(OH)(2)D(3))].We observed significant changes in RNAs of transcription factors, leptin, peptide hormones, and RNAs encoding proteins of fatty acid, amino acid, xenobiotic metabolism, receptor-activator of NFκB ligand (RANKL), and calcitonin-like ligand receptor pathways. Early highly restricted, and subsequent massive changes in more than 10% of expressed cellular RNA were observed. At days post fertilization (dpf) 2 [24 h 1α,25(OH)(2)D(3)-treatment], only four RNAs were differentially expressed (hormone vs. vehicle). On dpf 4 (72 h treatment), 77 RNAs; on dpf 6 (120 h treatment) 1039 RNAs; and on dpf 7 (144 h treatment), 2407 RNAs were differentially expressed in response to 1α,25(OH)(2)D(3). Fewer RNAs (n = 481) were altered in dpf 7 larvae treated for 24 h with 1α,25(OH)(2)D(3) vs. those treated with hormone for 144 h. At dpf 7, in 1α,25(OH)(2)D(3)-treated larvae, pharyngeal cartilage was larger and mineralization was greater. Changes in expression of RNAs for transcription factors, peptide hormones, and RNAs encoding proteins integral to fatty acid, amino acid, leptin, calcitonin-like ligand receptor, RANKL, and xenobiotic metabolism pathways, demonstrate heretofore unrecognized mechanisms by which 1α,25(OH)(2)D(3) functions in vivo in developing eukaryotes.
Figures
References
-
- DeLuca HF. 2004. Overview of general physiologic features and functions of vitamin D. Am J Clin Nutr 80:1689S–1696S - PubMed
-
- Kumar R. 1991. Vitamin D and calcium transport. Kidney Int 40:1177–1189 - PubMed
-
- Haussler MR, Haussler CA, Bartik L, Whitfield GK, Hsieh JC, Slater S, Jurutka PW. 2008. Vitamin D receptor: molecular signaling and actions of nutritional ligands in disease prevention. Nutr Rev 66:S98–S112 - PubMed
-
- Wasserman RH, Smith CA, Brindak ME, De Talamoni N, Fullmer CS, Penniston JT, Kumar R. 1992. Vitamin D and mineral deficiencies increase the plasma membrane calcium pump of chicken intestine. Gastroenterology 102:886–894 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
