

A kinetic Zipper model and the assembly of tobacco mosaic virus
- PMID: 22735535
- PMCID: PMC3379025
- DOI: 10.1016/j.bpj.2012.05.007
A kinetic Zipper model and the assembly of tobacco mosaic virus
Abstract
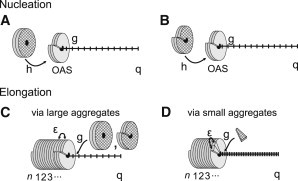
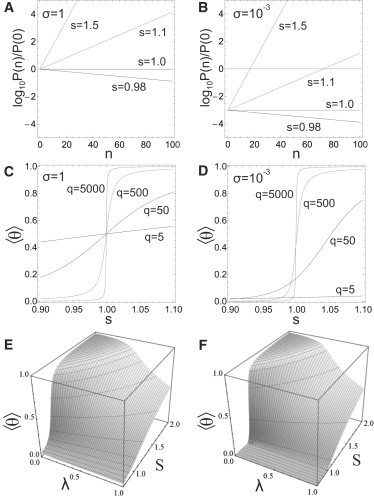
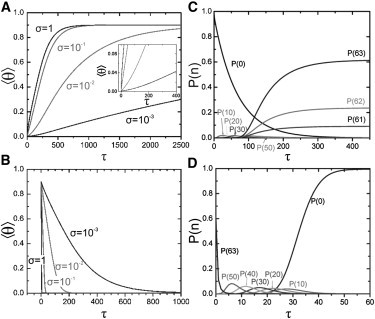
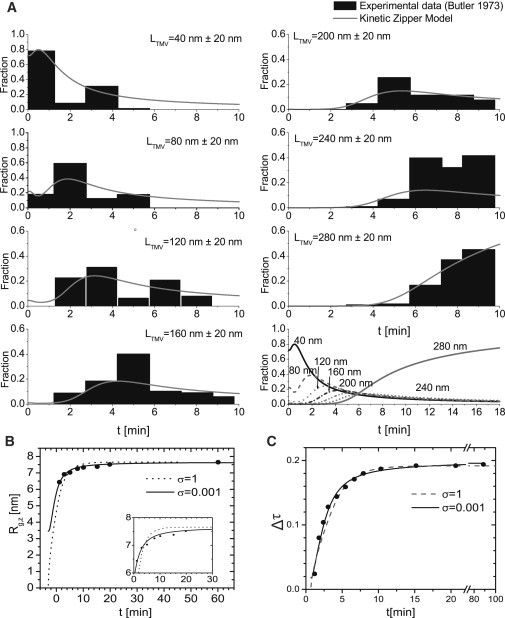
We put forward a modified Zipper model inspired by the statics and dynamics of the spontaneous reconstitution of rodlike tobacco mosaic virus particles in solutions containing the coat protein and the single-stranded RNA of the virus. An important ingredient of our model is an allosteric switch associated with the binding of the first protein unit to the origin-of-assembly domain of the viral RNA. The subsequent addition and conformational switching of coat proteins to the growing capsid we believe is catalyzed by the presence of the helical arrangement of bound proteins to the RNA. The model explains why the formation of complete viruses is favored over incomplete ones, even though the process is quasi-one-dimensional in character. We numerically solve the relevant kinetic equations and show that time evolution is different for the assembly and disassembly of the virus, the former exhibiting a time lag even if all forward rate constants are equal. We find the late-stage assembly kinetics in the presence of excess protein to be governed by a single-exponential relaxation, which agrees with available experimental data on TMV reconstruction.
Copyright © 2012 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Elucidation of the viral disassembly switch of tobacco mosaic virus.EMBO Rep. 2019 Nov 5;20(11):e48451. doi: 10.15252/embr.201948451. Epub 2019 Sep 19. EMBO Rep. 2019. PMID: 31535454 Free PMC article.
-
Hydrogen-bonding networks and RNA bases revealed by cryo electron microscopy suggest a triggering mechanism for calcium switches.Proc Natl Acad Sci U S A. 2011 Jun 7;108(23):9637-42. doi: 10.1073/pnas.1018104108. Epub 2011 May 17. Proc Natl Acad Sci U S A. 2011. PMID: 21586634 Free PMC article.
-
Coat protein interactions involved in tobacco mosaic tobamovirus cross-protection.Virology. 1998 Sep 1;248(2):188-98. doi: 10.1006/viro.1998.9280. Virology. 1998. PMID: 9721228
-
Satellite tobacco mosaic virus RNA: structure and implications for assembly.Curr Opin Struct Biol. 2001 Feb;11(1):59-65. doi: 10.1016/s0959-440x(00)00166-4. Curr Opin Struct Biol. 2001. PMID: 11179893 Review.
-
Tobacco mosaic virus and the study of early events in virus infections.Philos Trans R Soc Lond B Biol Sci. 1999 Mar 29;354(1383):603-11. doi: 10.1098/rstb.1999.0412. Philos Trans R Soc Lond B Biol Sci. 1999. PMID: 10212940 Free PMC article. Review.
Cited by
-
Design and self-assembly of simple coat proteins for artificial viruses.Nat Nanotechnol. 2014 Sep;9(9):698-702. doi: 10.1038/nnano.2014.169. Epub 2014 Aug 24. Nat Nanotechnol. 2014. PMID: 25150720
-
A kinetic model for the impact of packaging signal mimics on genome encapsulation.Biophys J. 2022 Jul 5;121(13):2583-2599. doi: 10.1016/j.bpj.2022.05.040. Epub 2022 May 30. Biophys J. 2022. PMID: 35642255 Free PMC article.
-
Hyperstretching DNA.Nat Commun. 2017 Dec 19;8(1):2197. doi: 10.1038/s41467-017-02396-1. Nat Commun. 2017. PMID: 29259297 Free PMC article.
-
Disulfide Reduction Allosterically Destabilizes the β-Ladder Subdomain Assembly within the NS1 Dimer of ZIKV.Biophys J. 2020 Oct 20;119(8):1525-1537. doi: 10.1016/j.bpj.2020.08.036. Epub 2020 Sep 6. Biophys J. 2020. PMID: 32946768 Free PMC article.
-
CRISPR-Guided Programmable Self-Assembly of Artificial Virus-Like Nucleocapsids.Nano Lett. 2021 Apr 14;21(7):2752-2757. doi: 10.1021/acs.nanolett.0c04640. Epub 2021 Mar 17. Nano Lett. 2021. PMID: 33729813 Free PMC article.
References
-
- Beijerinck M. About a contagium vivum fluidum as the cause of the mottling of tobacco leaves [Over een contagium vivum fluidum als oorzaak van de vlekziekte der tabaksbladen] Verhandelingen der Koninklijke Nederlandse Akademie van Wetenschappen. 1898;65:3–21.
-
- Scholthof K., Shaw J., Sindelar L. APS Press; St. Paul, MN: 1999. Tobacco Mosaic Virus: One Hundred Years of Contributions to Virology.
-
- Scholthof K.B. Tobacco mosaic virus: a model system for plant biology. Annu. Rev. Phytopathol. 2004;42:13–34. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
