

piRNAs can trigger a multigenerational epigenetic memory in the germline of C. elegans
- PMID: 22738725
- PMCID: PMC3464430
- DOI: 10.1016/j.cell.2012.06.018
piRNAs can trigger a multigenerational epigenetic memory in the germline of C. elegans
Abstract
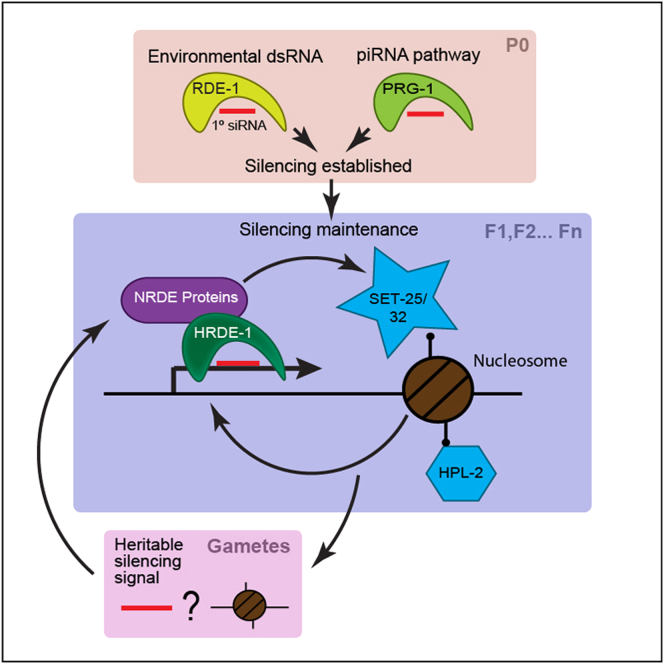
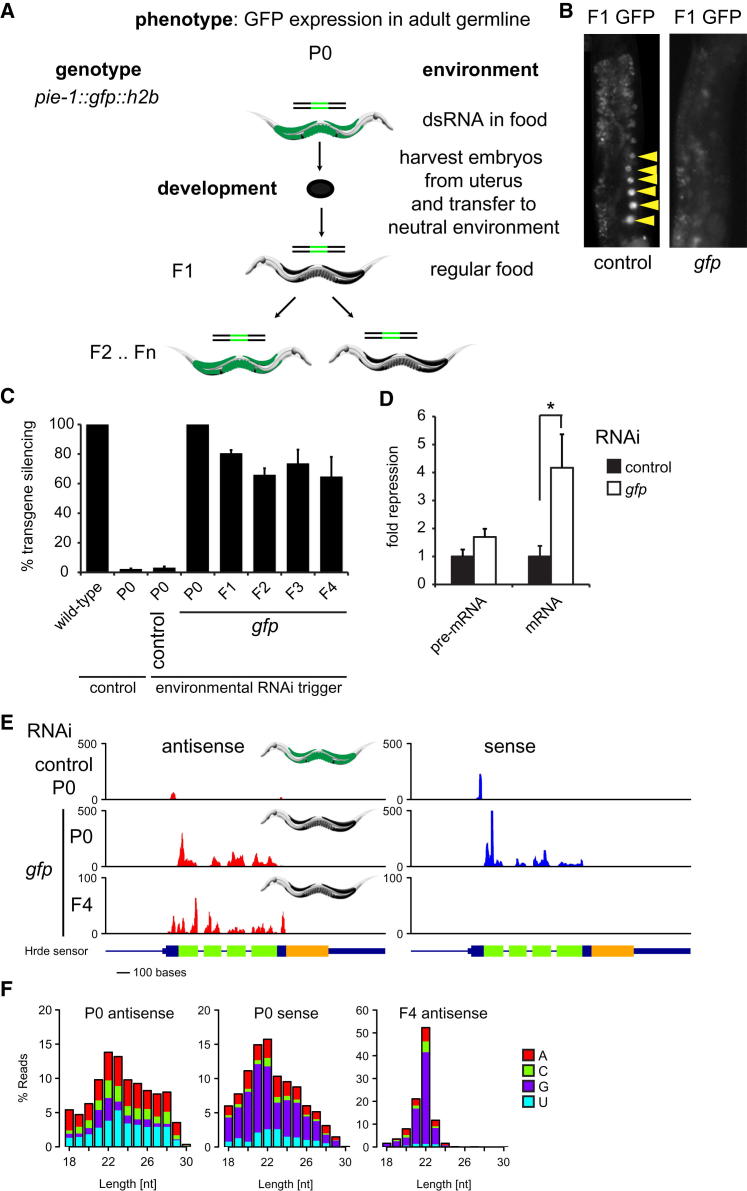
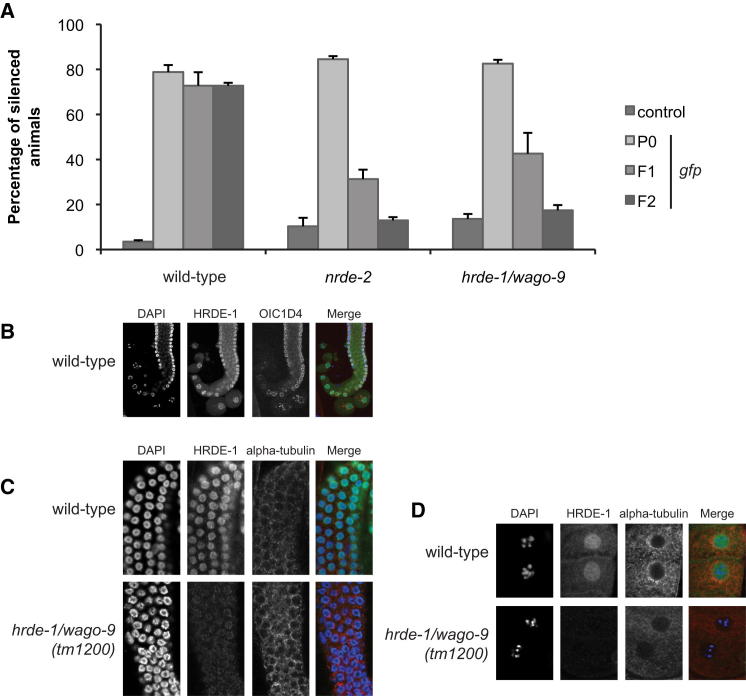
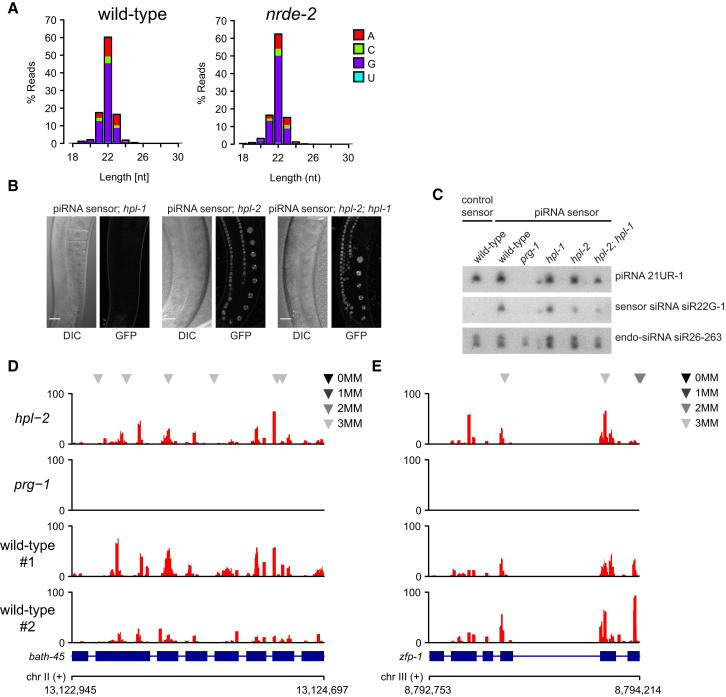
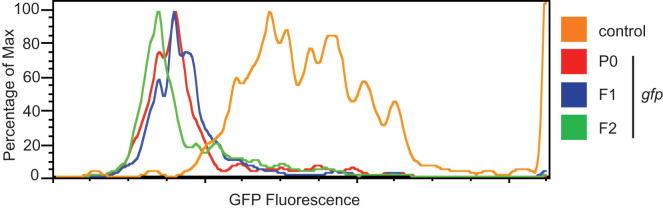
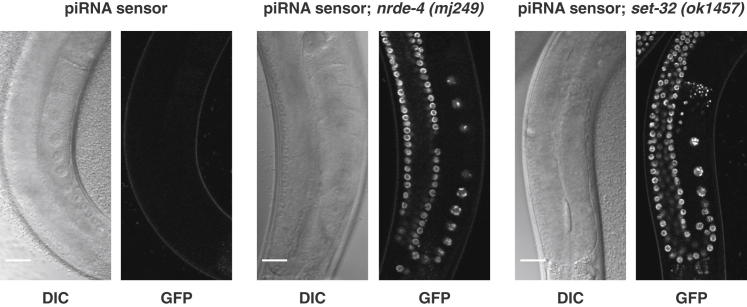
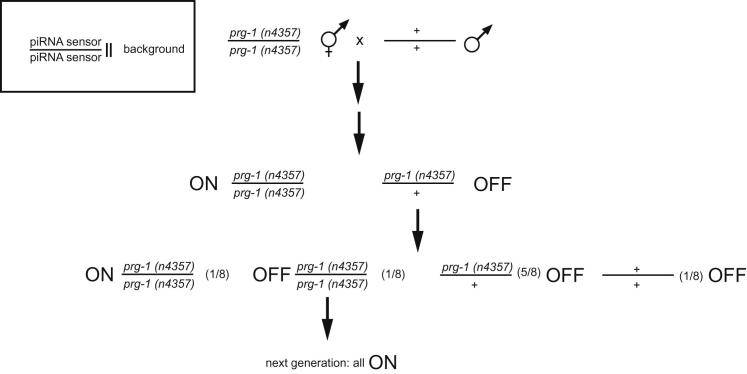
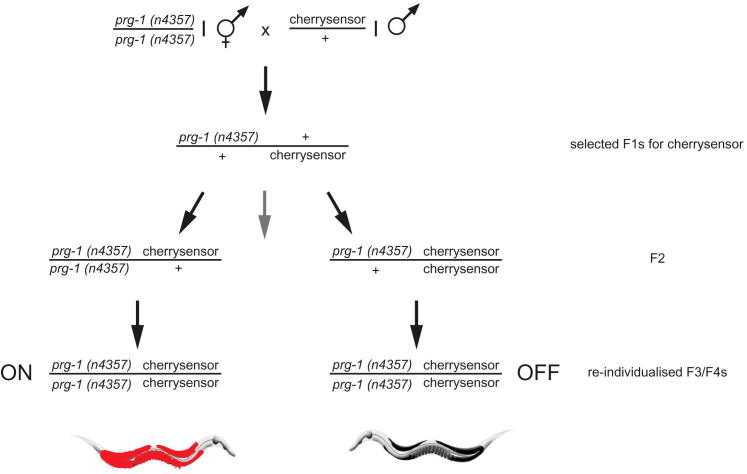
Transgenerational effects have wide-ranging implications for human health, biological adaptation, and evolution; however, their mechanisms and biology remain poorly understood. Here, we demonstrate that a germline nuclear small RNA/chromatin pathway can maintain stable inheritance for many generations when triggered by a piRNA-dependent foreign RNA response in C. elegans. Using forward genetic screens and candidate approaches, we find that a core set of nuclear RNAi and chromatin factors is required for multigenerational inheritance of environmental RNAi and piRNA silencing. These include a germline-specific nuclear Argonaute HRDE1/WAGO-9, a HP1 ortholog HPL-2, and two putative histone methyltransferases, SET-25 and SET-32. piRNAs can trigger highly stable long-term silencing lasting at least 20 generations. Once established, this long-term memory becomes independent of the piRNA trigger but remains dependent on the nuclear RNAi/chromatin pathway. Our data present a multigenerational epigenetic inheritance mechanism induced by piRNAs.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
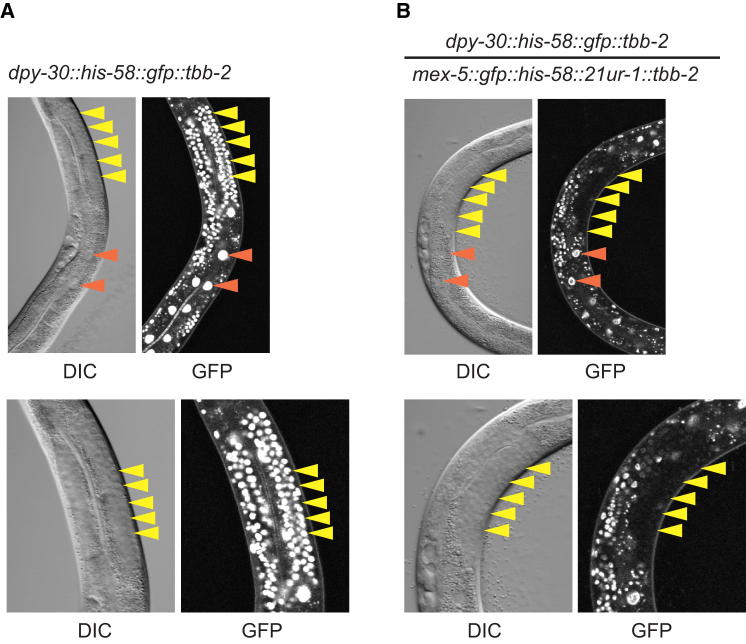
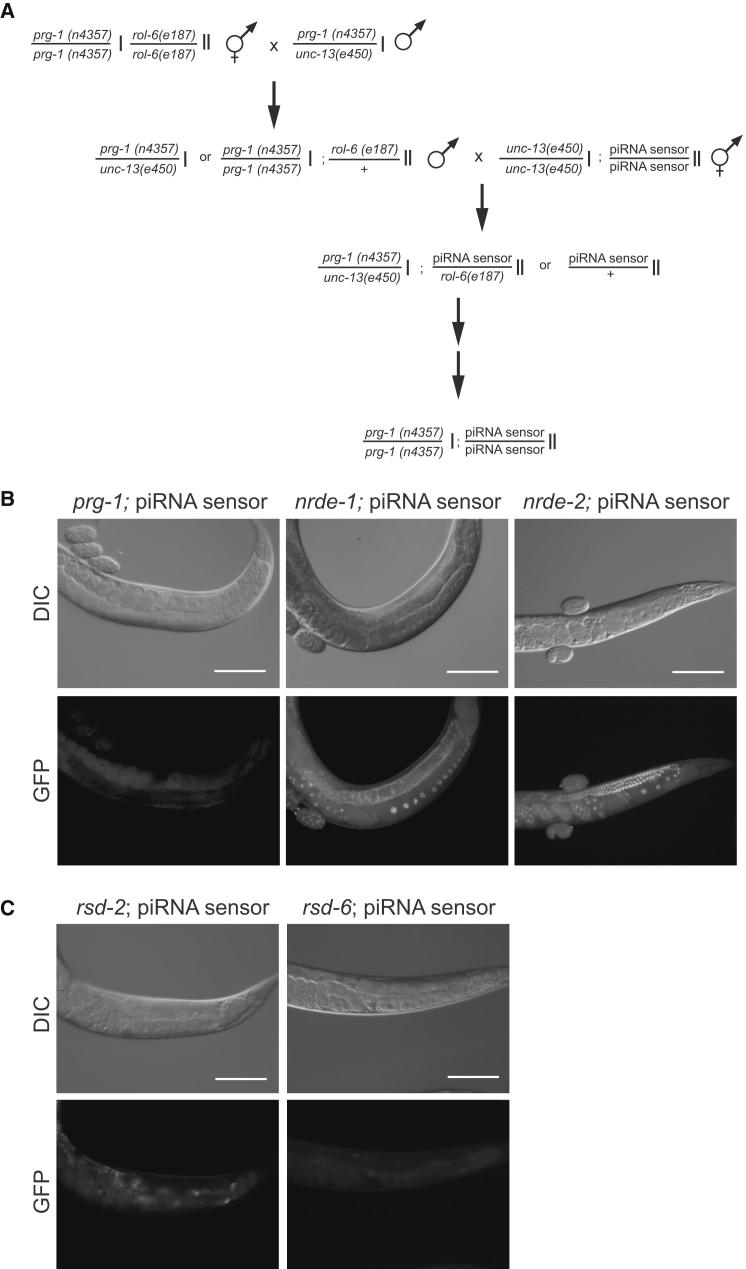
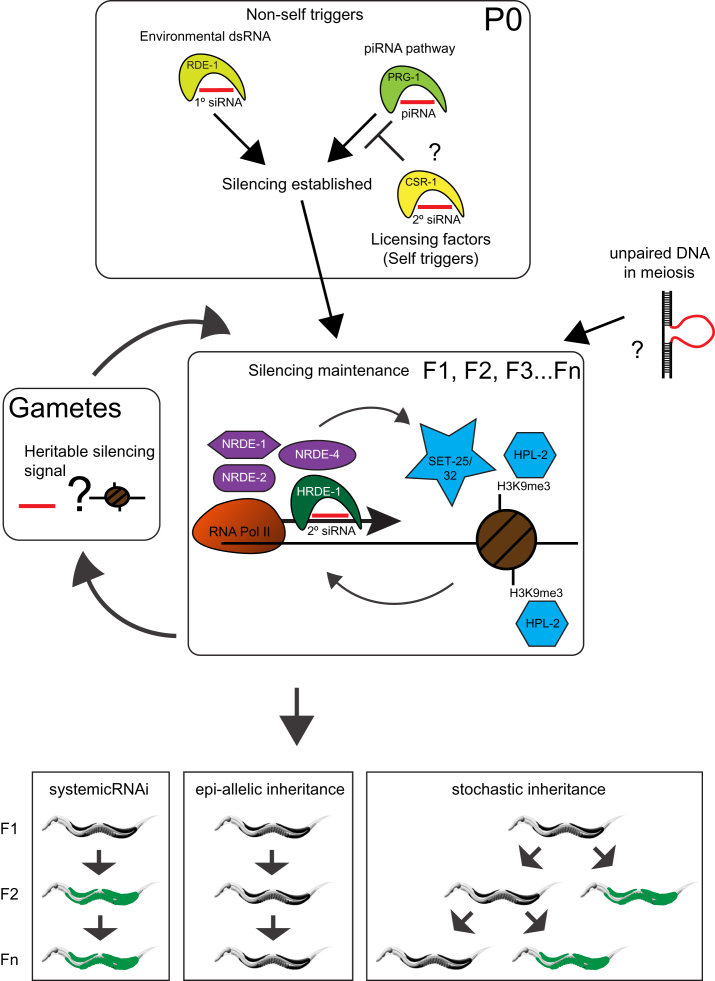
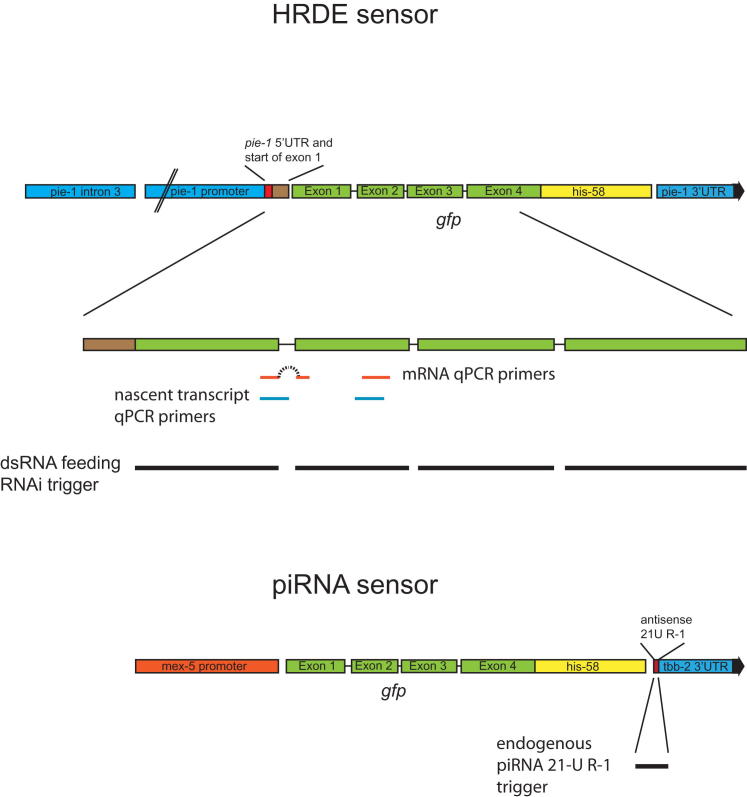
Comment in
-
Small RNAs: piRNA surveillance in the C. elegans germline.Nat Rev Genet. 2012 Jul 10;13(8):518-9. doi: 10.1038/nrg3289. Nat Rev Genet. 2012. PMID: 22777129 No abstract available.
-
Small RNAs: transmitting silence through generations.Nat Rev Mol Cell Biol. 2012 Jul 23;13(8):477. doi: 10.1038/nrm3406. Nat Rev Mol Cell Biol. 2012. PMID: 22820881 No abstract available.
References
-
- Bateson W. Dr. Kammerer's Testimony to the Inheritance of Acquired Characters. Nature. 1919;103:344–345.
Supplemental References
-
- Bruinsma, J.J., Schneider, D.L., Davis, D.E., and Kornfeld, K. (2008). Identification of mutations in Caenorhabditis elegans that cause resistance to high levels of dietary zinc and analysis using a genomewide map of single nucleotide polymorphisms scored by pyrosequencing. Genetics 179, 811–828. - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
