

Memory CD8+ T cells specific for a single immunodominant or subdominant determinant induced by peptide-dendritic cell immunization protect from an acute lethal viral disease
- PMID: 22740418
- PMCID: PMC3446599
- DOI: 10.1128/JVI.00981-12
Memory CD8+ T cells specific for a single immunodominant or subdominant determinant induced by peptide-dendritic cell immunization protect from an acute lethal viral disease
Abstract
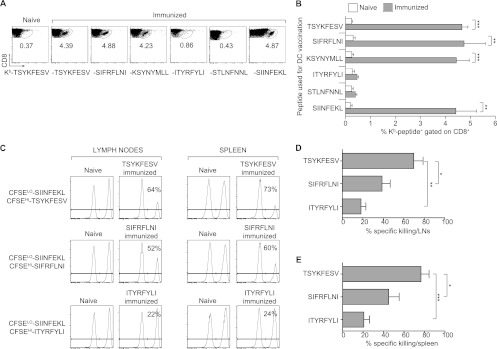
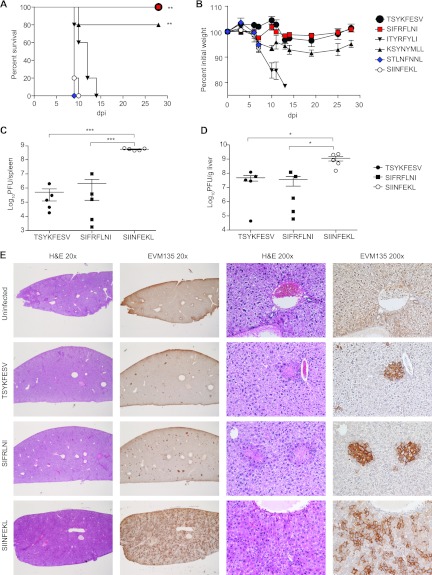
The antigens recognized by individual CD8(+) T cells are small peptides bound to major histocompatibility complex (MHC) class I molecules. The CD8(+) T cell response to a virus is restricted to several peptides, and the magnitudes of the effector as well as memory phases of the response to the individual peptides are generally hierarchical. The peptide eliciting a stronger response is called immunodominant (ID), and those with smaller-magnitude responses are termed subdominant (SD). The relative importance of ID and SD determinants in protective immunity remains to be fully elucidated. We previously showed that multispecific memory CD8(+) T cells can protect susceptible mice from mousepox, an acute lethal viral disease. It remained unknown, however, whether CD8(+) T cells specific for single ID or SD peptides could be protective. Here, we demonstrate that immunization with dendritic cells pulsed with ID and some but not all SD peptides induces memory CD8(+) T cells that are fully capable of protecting susceptible mice from mousepox. Additionally, while natural killer (NK) cells are essential for the natural resistance of nonimmune C57BL/6 (B6) to mousepox, we show that memory CD8(+) T cells of single specificity also protect B6 mice depleted of NK cells. This suggests it is feasible to produce effective antiviral CD8(+) T cell vaccines using single CD8(+) T cell determinants and that NK cells are no longer essential when memory CD8(+) T cells are present.
Figures




Similar articles
-
Lipid-nanoparticle-encapsulated mRNA vaccines induce protective memory CD8 T cells against a lethal viral infection.Mol Ther. 2021 Sep 1;29(9):2769-2781. doi: 10.1016/j.ymthe.2021.05.011. Epub 2021 May 14. Mol Ther. 2021. PMID: 33992803 Free PMC article.
-
Loss of Resistance to Mousepox during Chronic Lymphocytic Choriomeningitis Virus Infection Is Associated with Impaired T-Cell Responses and Can Be Rescued by Immunization.J Virol. 2020 Feb 14;94(5):e01832-19. doi: 10.1128/JVI.01832-19. Print 2020 Feb 14. J Virol. 2020. PMID: 31826990 Free PMC article.
-
Direct CD28 costimulation is required for CD8+ T cell-mediated resistance to an acute viral disease in a natural host.J Immunol. 2006 Dec 1;177(11):8027-36. doi: 10.4049/jimmunol.177.11.8027. J Immunol. 2006. PMID: 17114476
-
Heterotypic immunity against vaccinia virus in an HLA-B*07:02 transgenic mousepox infection model.Sci Rep. 2020 Aug 5;10(1):13167. doi: 10.1038/s41598-020-69897-w. Sci Rep. 2020. PMID: 32759969 Free PMC article.
-
The Pathogenesis and Immunobiology of Mousepox.Adv Immunol. 2016;129:251-76. doi: 10.1016/bs.ai.2015.10.001. Epub 2015 Nov 21. Adv Immunol. 2016. PMID: 26791861 Review.
Cited by
-
Asymptomatic HLA-A*02:01-restricted epitopes from herpes simplex virus glycoprotein B preferentially recall polyfunctional CD8+ T cells from seropositive asymptomatic individuals and protect HLA transgenic mice against ocular herpes.J Immunol. 2013 Nov 15;191(10):5124-38. doi: 10.4049/jimmunol.1301415. Epub 2013 Oct 7. J Immunol. 2013. PMID: 24101547 Free PMC article.
-
Sizing up the key determinants of the CD8(+) T cell response.Nat Rev Immunol. 2015 Nov;15(11):705-16. doi: 10.1038/nri3905. Epub 2015 Oct 9. Nat Rev Immunol. 2015. PMID: 26449178 Review.
-
Discovering protective CD8 T cell epitopes--no single immunologic property predicts it!Curr Opin Immunol. 2015 Jun;34:43-51. doi: 10.1016/j.coi.2015.01.013. Epub 2015 Feb 6. Curr Opin Immunol. 2015. PMID: 25660347 Free PMC article. Review.
-
Memory CD8⁺ T cells can outsource IFN-γ production but not cytolytic killing for antiviral protection.Cell Host Microbe. 2013 May 15;13(5):546-557. doi: 10.1016/j.chom.2013.04.004. Cell Host Microbe. 2013. PMID: 23684306 Free PMC article.
-
Modification of a tumor antigen determinant to improve peptide/MHC stability is associated with increased immunogenicity and cross-priming a larger fraction of CD8+ T cells.J Immunol. 2012 Dec 15;189(12):5549-60. doi: 10.4049/jimmunol.1102221. Epub 2012 Nov 21. J Immunol. 2012. PMID: 23175697 Free PMC article.
References
-
- Abbas AK. 1989. Antigen presentation by B lymphocytes: mechanisms and functional significance. Semin. Immunol. 1:5–12 - PubMed
-
- Amoscato AA, Prenovitz DA, Lotze MT. 1998. Rapid extracellular degradation of synthetic class I peptides by human dendritic cells. J. Immunol. 161:4023–4032 - PubMed
-
- Badovinac VP, Messingham KA, Jabbari A, Haring JS, Harty JT. 2005. Accelerated CD8+ T-cell memory and prime-boost response after dendritic-cell vaccination. Nat. Med. 11:748–756 - PubMed
-
- Bolitho P, Voskoboinik I, Trapani JA, Smyth MJ. 2007. Apoptosis induced by the lymphocyte effector molecule perforin. Curr. Opin. Immunol. 19:339–347 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
