

Identification of cytoskeletal elements enclosing the ATP pools that fuel human red blood cell membrane cation pumps
- PMID: 22745158
- PMCID: PMC3412019
- DOI: 10.1073/pnas.1209014109
Identification of cytoskeletal elements enclosing the ATP pools that fuel human red blood cell membrane cation pumps
Abstract
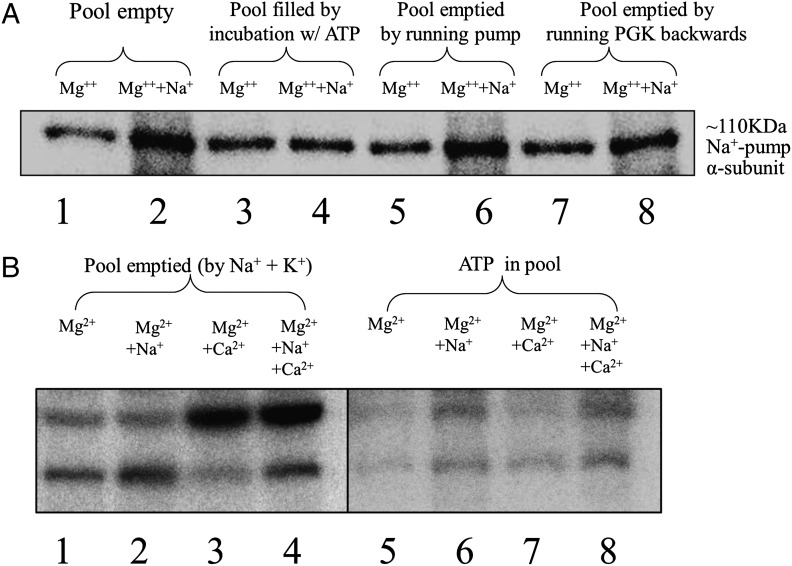
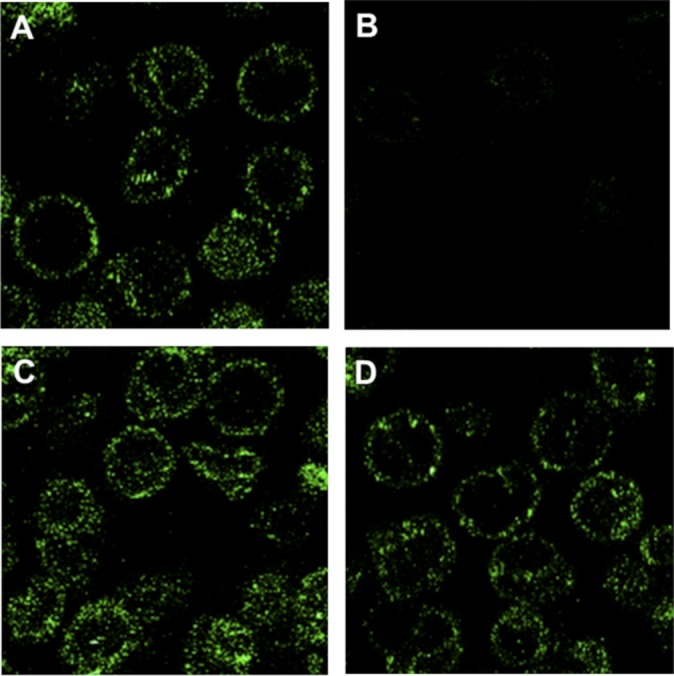
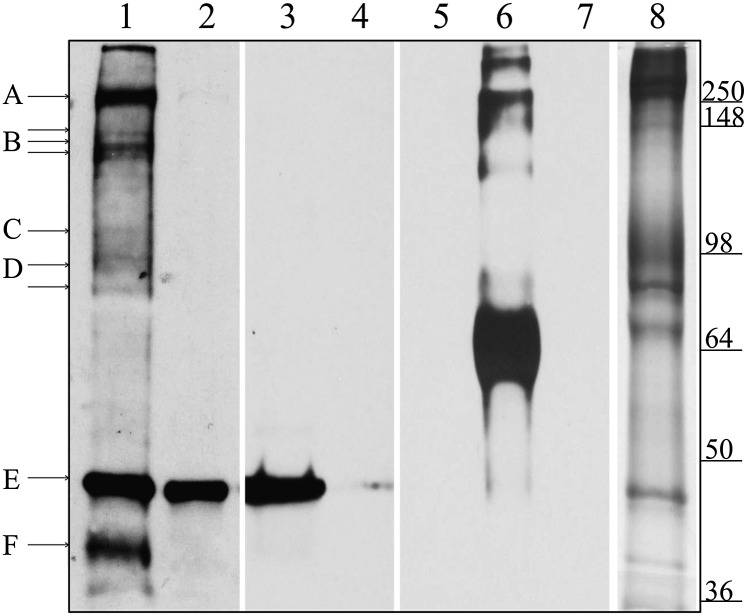
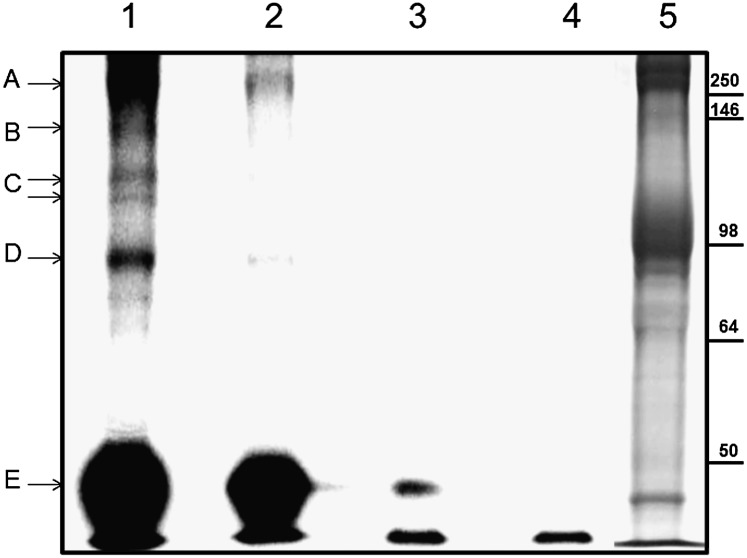
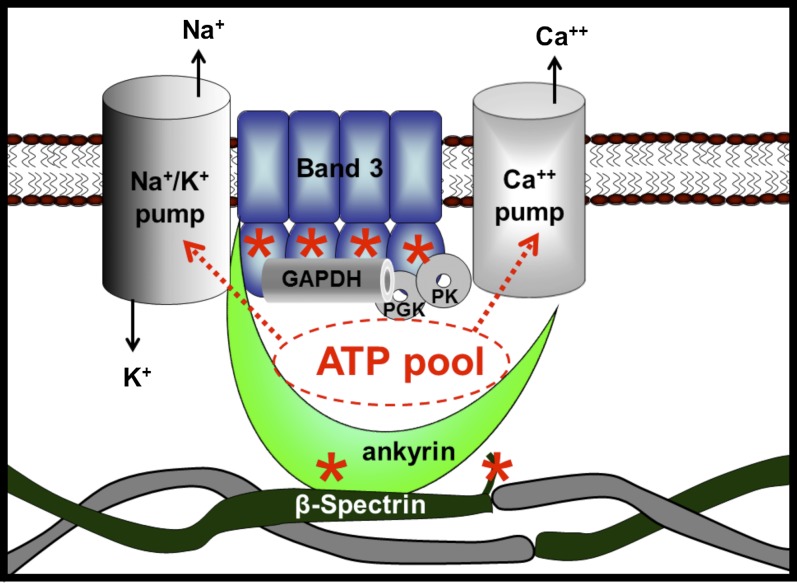
The type of metabolic compartmentalization that occurs in red blood cells differs from the types that exist in most eukaryotic cells, such as intracellular organelles. In red blood cells (ghosts), ATP is sequestered within the cytoskeletal-membrane complex. These pools of ATP are known to directly fuel both the Na(+)/K(+) and Ca(2+) pumps. ATP can be entrapped within these pools either by incubation with bulk ATP or by operation of the phosphoglycerate kinase and pyruvate kinase reactions to enzymatically generate ATP. When the pool is filled with nascent ATP, metabolic labeling of the Na(+)/K(+) or Ca(2+) pump phosphoproteins (E(Na)-P and E(Ca)-P, respectively) from bulk [γ-(32)P]-ATP is prevented until the pool is emptied by various means. Importantly, the pool also can be filled with the fluorescent ATP analog trinitrophenol ATP, as well as with a photoactivatable ATP analog, 8-azido-ATP (N(3)-ATP). Using the fluorescent ATP, we show that ATP accumulates and then disappears from the membrane as the ATP pools are filled and subsequently emptied, respectively. By loading N(3)-ATP into the membrane pool, we demonstrate that membrane proteins that contribute to the pool's architecture can be photolabeled. With the aid of an antibody to N(3)-ATP, we identify these labeled proteins by immunoblotting and characterize their derived peptides by mass spectrometry. These analyses show that the specific peptides that corral the entrapped ATP derive from sequences within β-spectrin, ankyrin, band 3, and GAPDH.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Alberts B, et al. In: Molecular Biology of the Cell. 4th Ed. Alberts B, et al., editors. New York: Garland Science; 2002. pp. 659–669.
-
- Srere PA. Complexes of sequential metabolic enzymes. Annu Rev Biochem. 1987;56:89–124. - PubMed
-
- Skou JC. The influence of some cations on an adenosine triphosphatase from peripheral nerves. Biochim Biophys Acta. 1957;23:394–401. - PubMed
-
- Schatzmann HJ. ATP-dependent Ca++-extrusion from human red cells. Experientia. 1966;22:364–365. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
