

β-Catenin signaling specifies progenitor cell identity in parallel with Shh signaling in the developing mammalian thalamus
- PMID: 22745311
- PMCID: PMC3392701
- DOI: 10.1242/dev.072314
β-Catenin signaling specifies progenitor cell identity in parallel with Shh signaling in the developing mammalian thalamus
Abstract
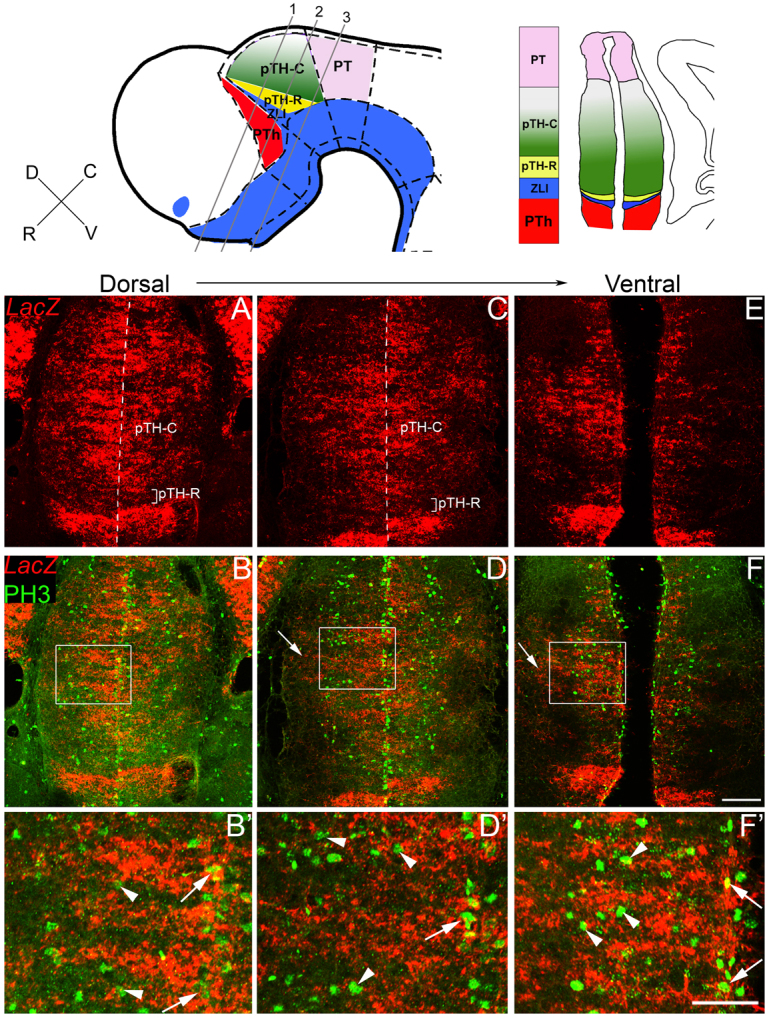
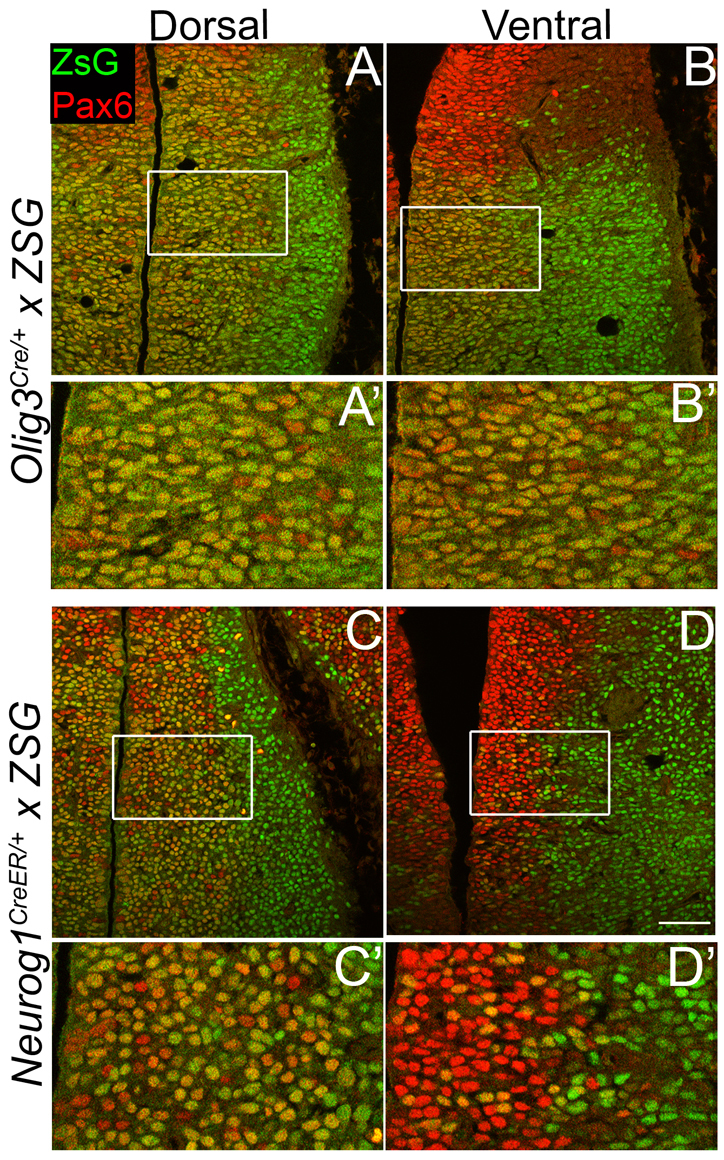
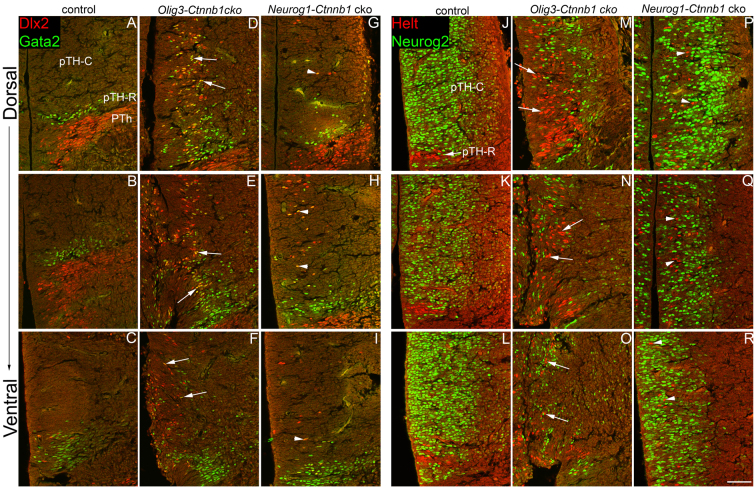
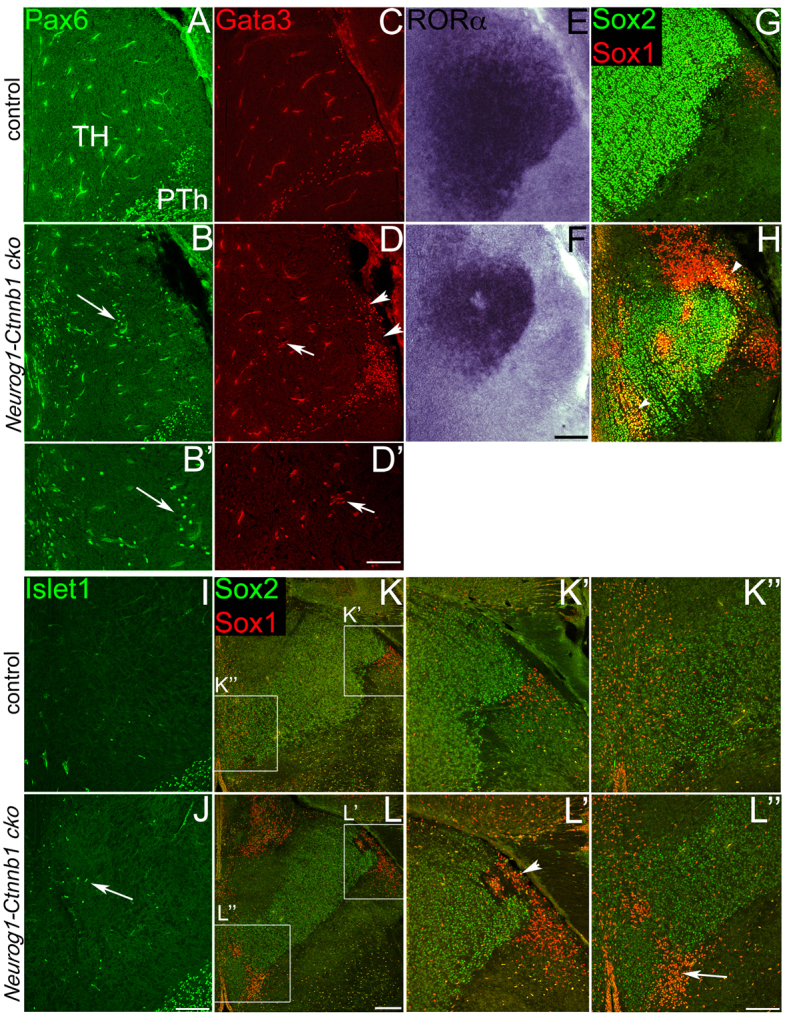
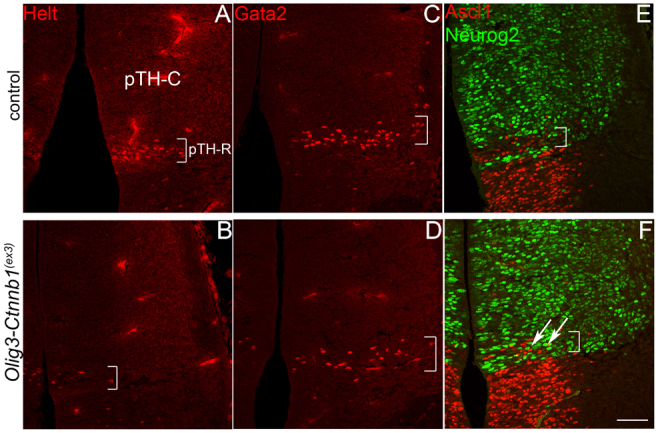
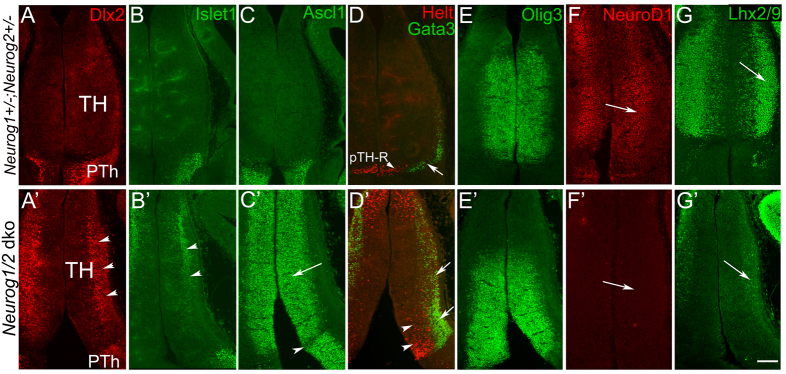
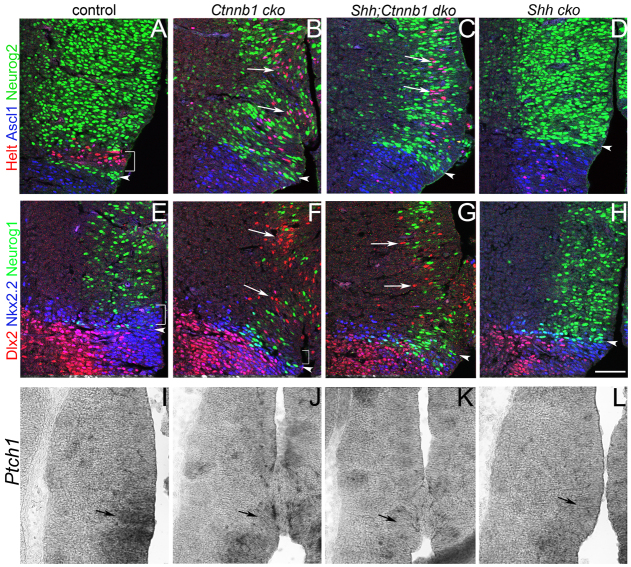
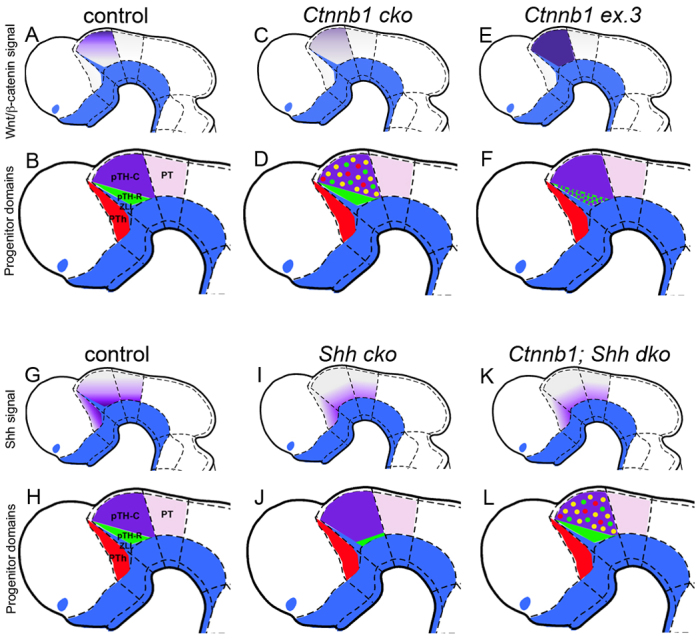
Neural progenitor cells within the developing thalamus are spatially organized into distinct populations. Their correct specification is critical for generating appropriate neuronal subtypes in specific locations during development. Secreted signaling molecules, such as sonic hedgehog (Shh) and Wnts, are required for the initial formation of the thalamic primordium. Once thalamic identity is established and neurogenesis is initiated, Shh regulates the positional identity of thalamic progenitor cells. Although Wnt/β-catenin signaling also has differential activity within the thalamus during this stage of development, its significance has not been directly addressed. In this study, we used conditional gene manipulations in mice and explored the roles of β-catenin signaling in the regional identity of thalamic progenitor cells. We found β-catenin is required during thalamic neurogenesis to maintain thalamic fate while suppressing prethalamic fate, demonstrating that regulation of regional fate continues to require extrinsic signals. These roles of β-catenin appeared to be mediated at least partly by regulating two basic helix-loop-helix (bHLH) transcription factors, Neurog1 and Neurog2. β-Catenin and Shh signaling function in parallel to specify two progenitor domains within the thalamus, where individual transcription factors expressed in each progenitor domain were regulated differently by the two signaling pathways. We conclude that β-catenin has multiple functions during thalamic neurogenesis and that both Shh and β-catenin pathways are important for specifying distinct types of thalamic progenitor cells, ensuring that the appropriate neuronal subtypes are generated in the correct locations.
Figures
References
-
- Agren M., Kogerman P., Kleman M. I., Wessling M., Toftgard R. (2004). Expression of the Ptch1 tumor suppressor gene is regulated by alternative promotors and a single functional Gli-binding site. Gene 330, 101-114 - PubMed
-
- Alvarez-Medina R., Cayuso J., Okubo T., Takada S., Marti E. (2008). Wnt canonical pathway restricts graded Shh/Gli patterning activity through the regulation of Gli3 expression. Development 135, 237-247 - PubMed
-
- Backman M., Machon O., Mygland L., van den Bout C. J., Zhong W., Taketo M. M., Krauss S. (2005). Effects of canonical Wnt signaling on dorso-ventral specification of the mouse telencephalon. Dev. Biol. 279, 155-168 - PubMed
-
- Brault V., Moore R., Kutsch S., Ishibashi M., Rowitch D. H., McMahon A. P., Sommer L., Boussadia O., Kemler R. (2001). Inactivation of the b-catenin gene by Wnt1-Cre-mediated deletion results in dramatic brain malformation and failure of craniofacial development. Development 128, 1253-1264 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
