

Endothelial expression of TGFβ type II receptor is required to maintain vascular integrity during postnatal development of the central nervous system
- PMID: 22745736
- PMCID: PMC3383742
- DOI: 10.1371/journal.pone.0039336
Endothelial expression of TGFβ type II receptor is required to maintain vascular integrity during postnatal development of the central nervous system
Erratum in
- PLoS One. 2012 Sep;7(9). doi: 10.1371/annotation/8d859757-284b-406d-9cb9-a8776ad32fb1. McCarty, Joseph [corrected to McCarty, Joseph H] doi: 10.1371/annotation/8d859757-284b-406d-9cb9-a8776ad32fb1
Abstract
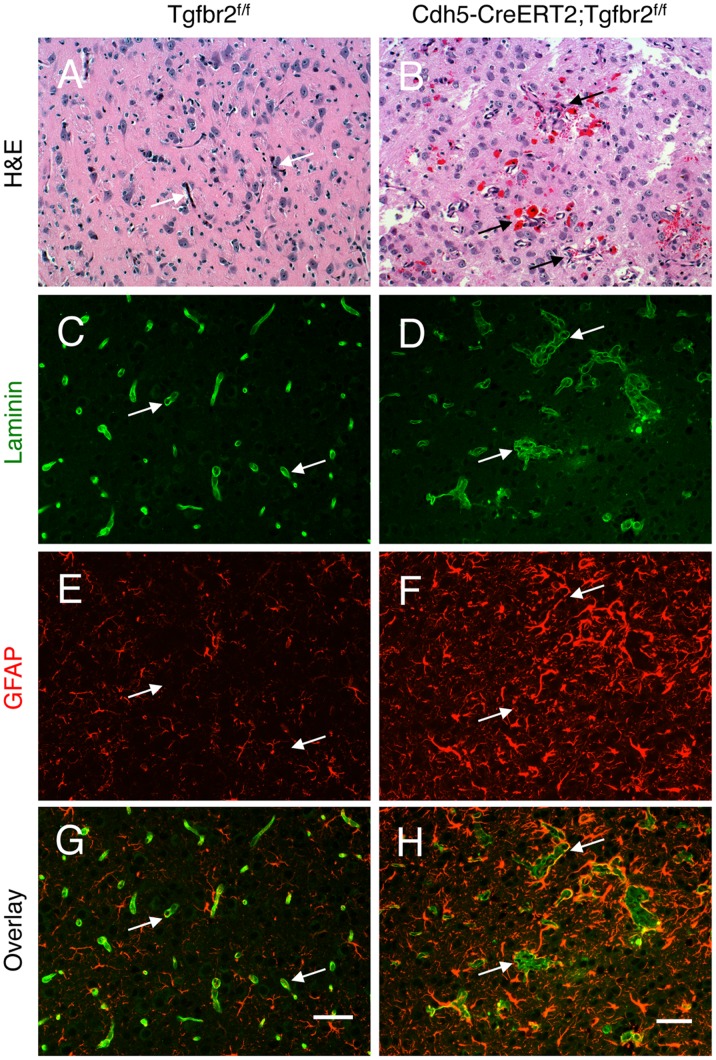
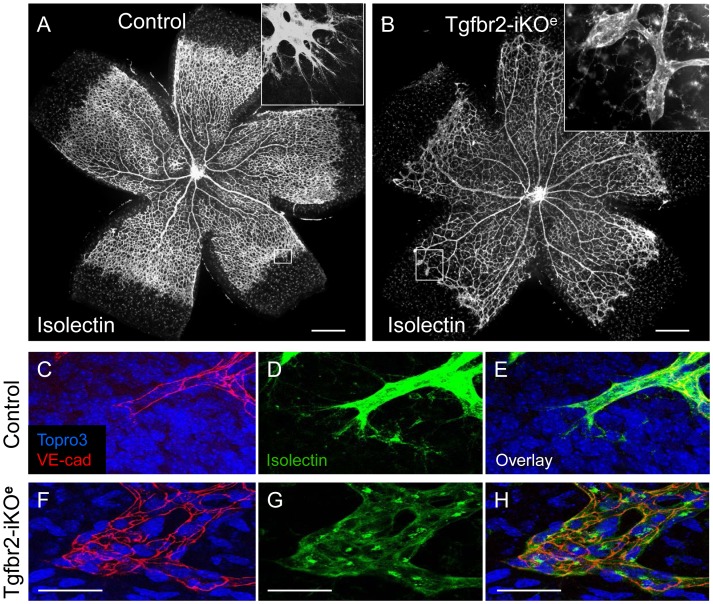
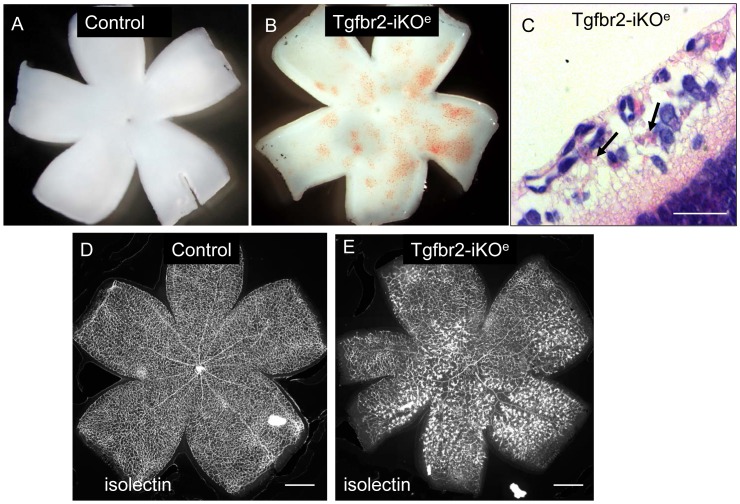
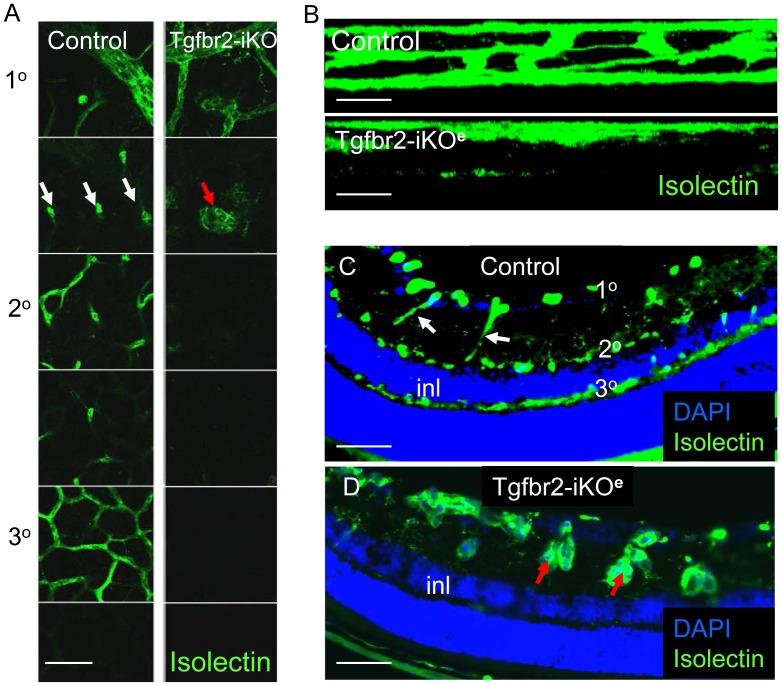
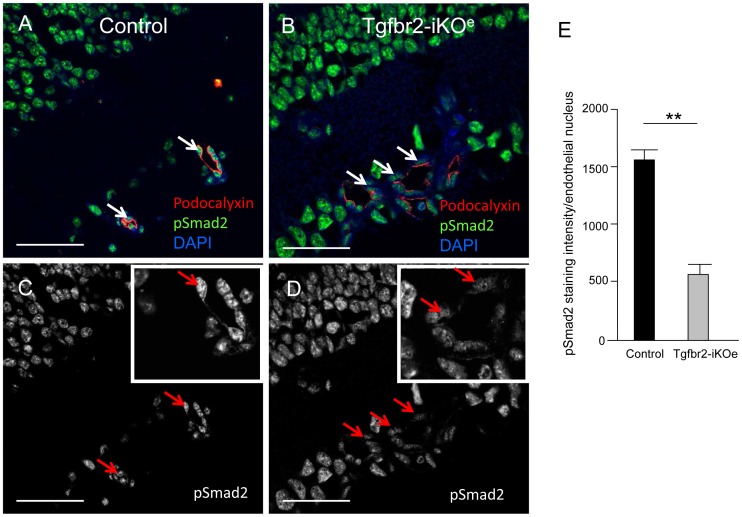
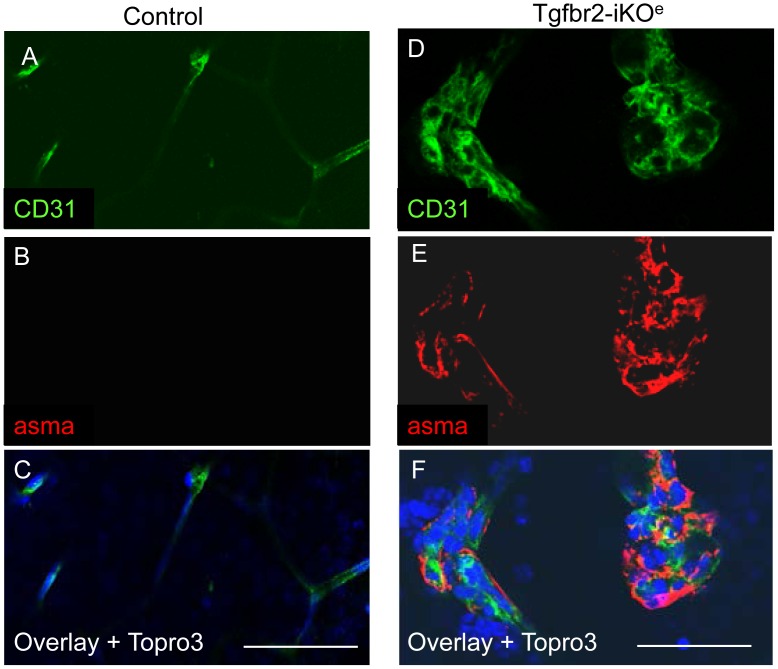
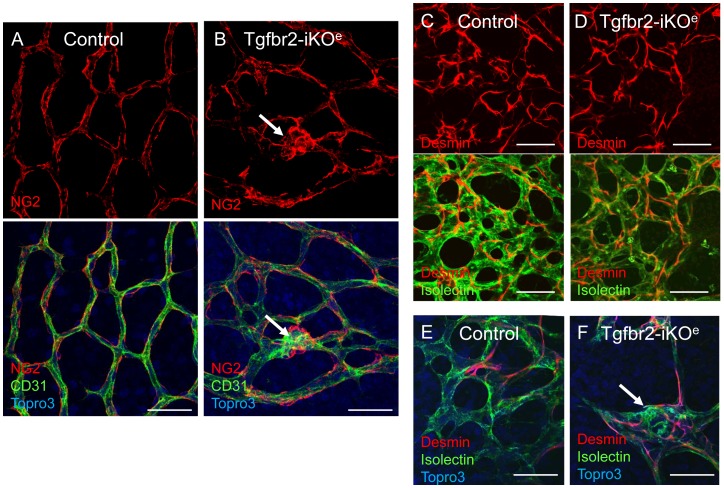
TGFβ signalling in endothelial cells is important for angiogenesis in early embryonic development, but little is known about its role in early postnatal life. To address this we used a tamoxifen inducible Cre-LoxP strategy in neonatal mice to deplete the TypeII TGFβ receptor (Tgfbr2) specifically in endothelial cells. This resulted in multiple micro-haemorrhages, and glomeruloid-like vascular tufts throughout the cerebral cortices and hypothalamus of the brain as well as in retinal tissues. A detailed examination of the retinal defects in these mutants revealed that endothelial adherens and tight junctions were in place, pericytes were recruited and there was no failure of vascular smooth muscle differentiation. However, the deeper retinal plexus failed to form in these mutants and the angiogenic sprouts stalled in their progress towards the inner nuclear layer. Instead the leading endothelial cells formed glomerular tufts with associated smooth muscle cells. This evidence suggests that TGFβ signalling is not required for vessel maturation, but is essential for the organised migration of endothelial cells as they begin to enter the deeper layers of the retina. Thus, TGFβ signalling is essential in vascular endothelial cells for maintaining vascular integrity at the angiogenic front as it migrates into developing neural tissues in early postnatal life.
Conflict of interest statement
Figures
References
-
- Armulik A, Genove G, Mae M, Nisancioglu MH, Wallgard E, et al. Pericytes regulate the blood-brain barrier. Nature. 2010;468:557–561. - PubMed
-
- Abbott NJ, Ronnback L, Hansson E. Astrocyte-endothelial interactions at the blood-brain barrier. Nat Rev Neurosci. 2006;7:41–53. - PubMed
-
- Ballabh P, Braun A, Nedergaard M. The blood-brain barrier: an overview: structure, regulation, and clinical implications. Neurobiol Dis. 2004;16:1–13. - PubMed
-
- Roland EH, Hill A. Germinal matrix-intraventricular hemorrhage in the premature newborn: management and outcome. Neurol Clin 21: 833–851, vi-vii. 2003. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
