

Inhibition of Eph receptor-ephrin ligand interaction by tea polyphenols
- PMID: 22750215
- PMCID: PMC3677025
- DOI: 10.1016/j.phrs.2012.05.010
Inhibition of Eph receptor-ephrin ligand interaction by tea polyphenols
Abstract
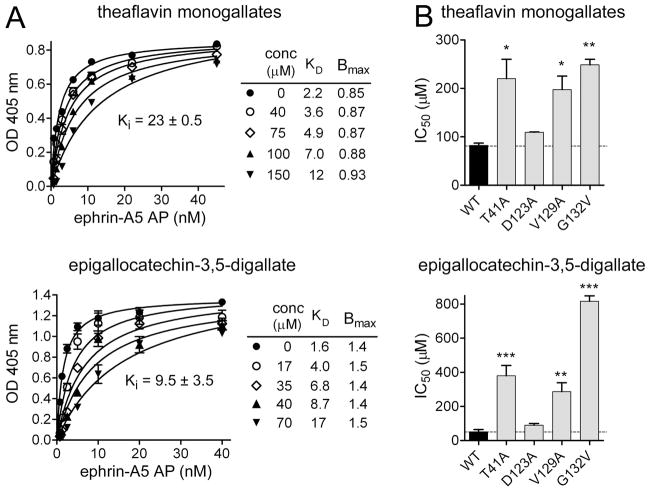
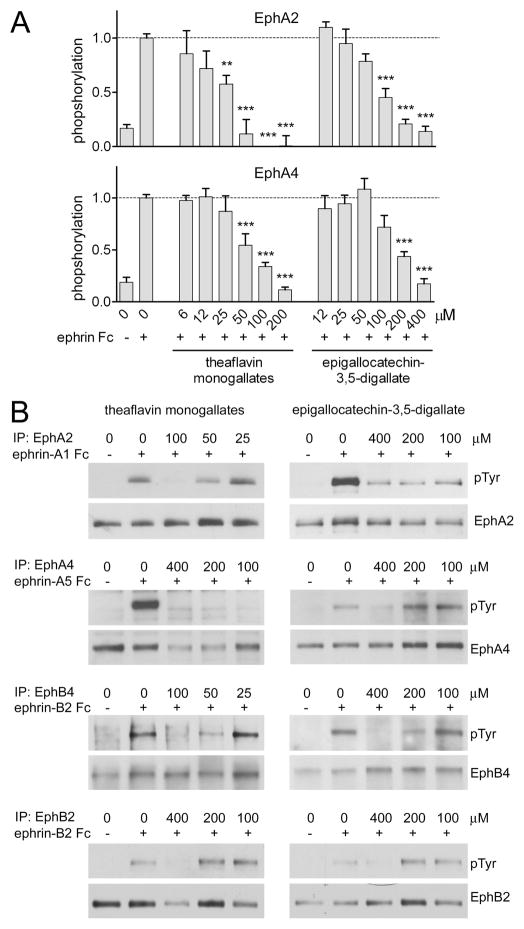
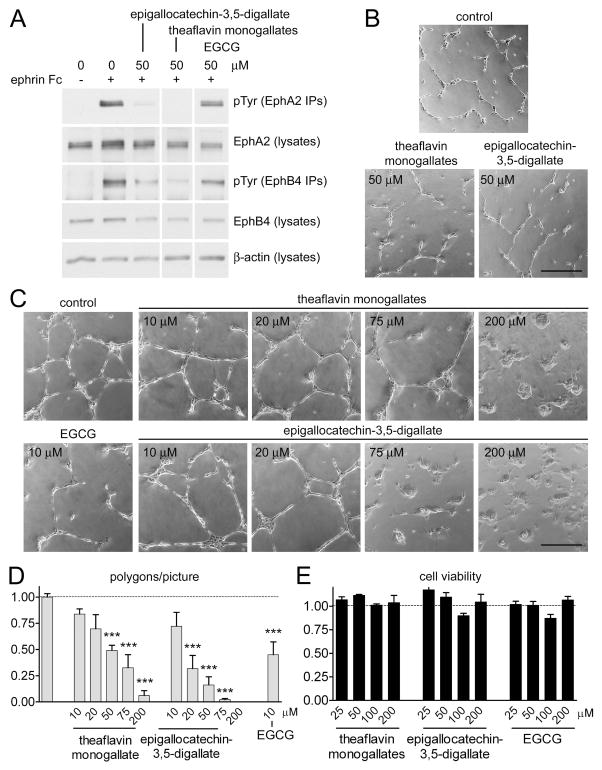
Tea contains a variety of bioactive chemicals, such as catechins and other polyphenols. These compounds are thought to be responsible for the health benefits of tea consumption by affecting the function of many cellular targets, not all of which have been identified. In a high-throughput screen for small molecule antagonists of the EphA4 receptor tyrosine kinase, we identified five tea polyphenols that substantially inhibit EphA4 binding to a synthetic peptide ligand. Further characterization of theaflavin monogallates from black tea and epigallocatechin-3,5-digallate from green tea revealed that these compounds at low micromolar concentrations also inhibit binding of the natural ephrin ligands to EphA4 and several other Eph receptors in in vitro assays. The compounds behave as competitive EphA4 antagonists, and their inhibitory activity is affected by amino acid mutations within the ephrin binding pocket of EphA4. In contrast, the major green tea catechin, epigallocatechin-3-gallate (EGCG), does not appear to be an effective Eph receptor antagonist. In cell culture assays, theaflavin monogallates and epigallocatechin-3,5-digallate inhibit ephrin-induced tyrosine phosphorylation (activation) of Eph receptors and endothelial capillary-like tube formation. However, the wider spectrum of Eph receptors affected by the tea derivatives in cells suggests additional mechanisms of inhibition besides interfering with ephrin binding. These results show that tea polyphenols derived from both black and green tea can suppress the biological activities of Eph receptors. Thus, the Eph receptor tyrosine kinase family represents an important class of targets for tea-derived phytochemicals.
Copyright © 2012 Elsevier Ltd. All rights reserved.
Figures

Similar articles
-
Small molecules can selectively inhibit ephrin binding to the EphA4 and EphA2 receptors.J Biol Chem. 2008 Oct 24;283(43):29461-72. doi: 10.1074/jbc.M804103200. Epub 2008 Aug 26. J Biol Chem. 2008. PMID: 18728010 Free PMC article.
-
A disalicylic acid-furanyl derivative inhibits ephrin binding to a subset of Eph receptors.Chem Biol Drug Des. 2011 Oct;78(4):667-78. doi: 10.1111/j.1747-0285.2011.01199.x. Epub 2011 Sep 6. Chem Biol Drug Des. 2011. PMID: 21791013 Free PMC article.
-
Suppression of extracellular signals and cell proliferation by the black tea polyphenol, theaflavin-3,3'-digallate.Carcinogenesis. 1999 Apr;20(4):733-6. doi: 10.1093/carcin/20.4.733. Carcinogenesis. 1999. PMID: 10223207
-
Eph Receptors as Drug Targets: Single-Chain Antibodies and Beyond.Curr Drug Targets. 2015;16(10):1021-30. doi: 10.2174/1389450116666150531154619. Curr Drug Targets. 2015. PMID: 26028047 Review.
-
Targeting the Eph System with Peptides and Peptide Conjugates.Curr Drug Targets. 2015;16(10):1031-47. doi: 10.2174/1389450116666150727115934. Curr Drug Targets. 2015. PMID: 26212263 Free PMC article. Review.
Cited by
-
An investigation of binding interactions of tumor-targeted peptide conjugated polyphenols with the kinase domain of ephrin B4 and B2 receptors.Mol Divers. 2024 Apr;28(2):817-849. doi: 10.1007/s11030-023-10621-x. Epub 2023 Feb 27. Mol Divers. 2024. PMID: 36847923 Free PMC article.
-
Role of Phytochemicals in Cancer Prevention.Int J Mol Sci. 2019 Oct 9;20(20):4981. doi: 10.3390/ijms20204981. Int J Mol Sci. 2019. PMID: 31600949 Free PMC article. Review.
-
Therapeutic potential of targeting the Eph/ephrin signaling complex.Int J Biochem Cell Biol. 2018 Dec;105:123-133. doi: 10.1016/j.biocel.2018.10.006. Epub 2018 Oct 19. Int J Biochem Cell Biol. 2018. PMID: 30343150 Free PMC article. Review.
-
Identification of Eph receptor signaling as a regulator of autophagy and a therapeutic target in colorectal carcinoma.Mol Oncol. 2019 Nov;13(11):2441-2459. doi: 10.1002/1878-0261.12576. Epub 2019 Oct 23. Mol Oncol. 2019. PMID: 31545551 Free PMC article.
-
Amino acid conjugates of lithocholic acid as antagonists of the EphA2 receptor.J Med Chem. 2013 Apr 11;56(7):2936-47. doi: 10.1021/jm301890k. Epub 2013 Mar 22. J Med Chem. 2013. PMID: 23489211 Free PMC article.
References
-
- Liao S, Kao YH, Hiipakka RA. Green tea: Biochemical and biological basis for health benefits. Vitam Horm. 2001;62:1–94. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
