

Metabolic differentiation in the embryonic retina
- PMID: 22750943
- PMCID: PMC3442239
- DOI: 10.1038/ncb2531
Metabolic differentiation in the embryonic retina
Abstract
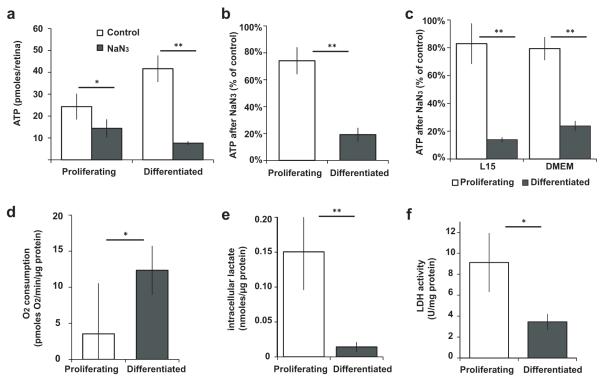
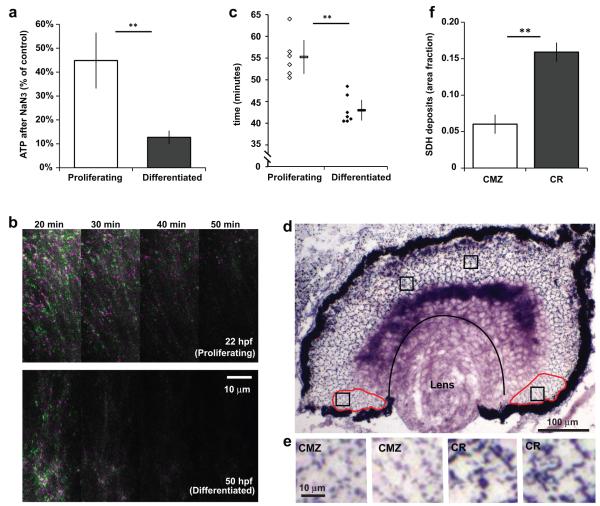
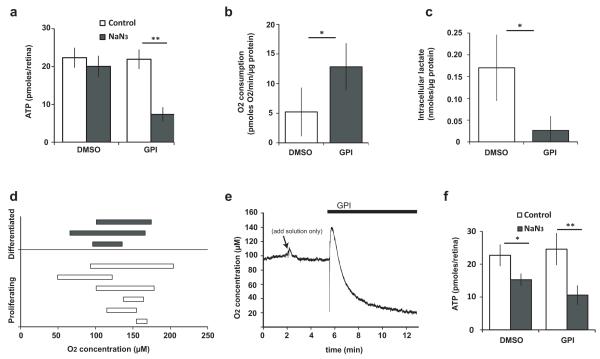
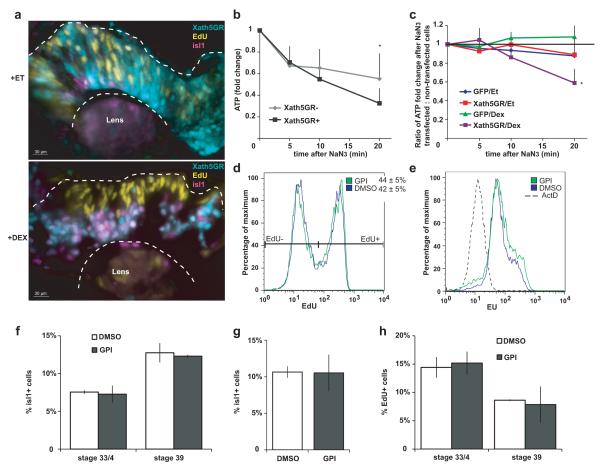
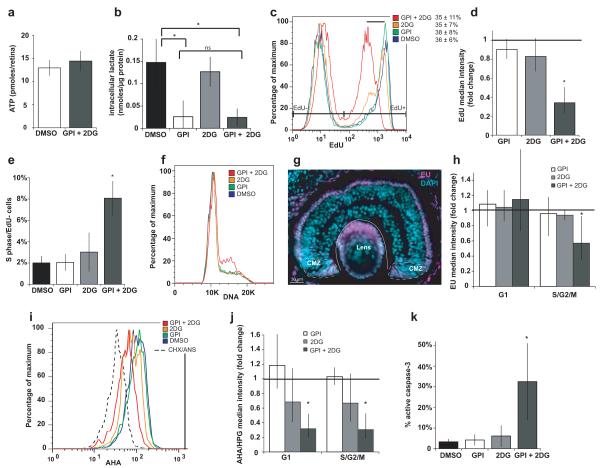
Unlike healthy adult tissues, cancers produce energy mainly by aerobic glycolysis instead of oxidative phosphorylation. This adaptation, called the Warburg effect, may be a feature of all dividing cells, both normal and cancerous, or it may be specific to cancers. It is not known whether, in a normally growing tissue during development, proliferating and postmitotic cells produce energy in fundamentally different ways. Here we show in the embryonic Xenopus retina in vivo, that dividing progenitor cells depend less on oxidative phosphorylation for ATP production than non-dividing differentiated cells, and instead use glycogen to fuel aerobic glycolysis. The transition from glycolysis to oxidative phosphorylation is connected to the cell differentiation process. Glycolysis is indispensable for progenitor proliferation and biosynthesis, even when it is not used for ATP production. These results suggest that the Warburg effect can be a feature of normal proliferation in vivo, and that the regulation of glycolysis and oxidative phosphorylation is critical for normal development.
Figures
References
-
- Warburg O. On the origin of cancer cells. Science. 1956;123:309–314. - PubMed
-
- Gatenby RA, Gillies RJ. Why do cancers have high aerobic glycolysis? Nat Rev Cancer. 2004;4:891–899. - PubMed
-
- Holt CE, Bertsch TW, Ellis HM, Harris WA. Cellular determination in the Xenopus retina is independent of lineage and birth date. Neuron. 1988;1:15–26. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
