

doi: 10.1101/cshperspect.a007922.
The evolution of the Wnt pathway
Affiliations
- PMID: 22751150
- PMCID: PMC3385961
- DOI: 10.1101/cshperspect.a007922
Item in Clipboard
The evolution of the Wnt pathway
Cold Spring Harb Perspect Biol.
.
Abstract
Wnt genes are important regulators of embryogenesis and cell differentiation in vertebrates and insects. New data revealed by comparative genomics have now shown that members of the Wnt signaling pathway can be found in all clades of metazoans, but not in fungi, plants, or unicellular eukaryotes. This article focuses on new data from recent genomic analyses of several basal metazoan organisms, providing evidence that the Wnt pathway was a primordial signaling pathway during evolution. The formation of a Wnt signaling center at the site of gastrulation was instrumental for the formation of a primary, anterior-posterior body axis, which can be traced throughout animal evolution.
Figures
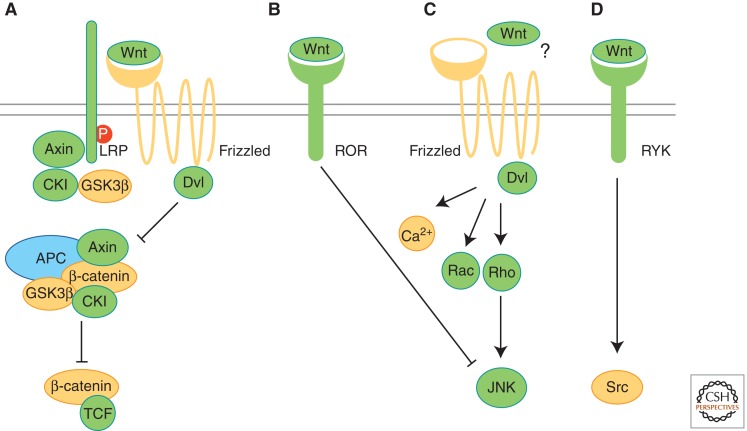
Evolution of Wnt pathways. Proteins involved in different Wnt pathways (A–D) are shown and labeled according to their occurrence in protozoans (yellow), pre-bilaterians (green), and bilaterians (blue). (A) Binding of Wnt to Frizzled receptor and LRP activates β-catenin/TCF signaling. The β-catenin destruction complex, including APC, Axin, GSK3β, and CKI, sequesters and phosphorylates β-catenin. Phosphorylation of LRP is crucial for Wnt/β-catenin signaling. (B) Binding of Wnt to the receptor tyrosine kinase (RTK) Ror2 activates Jnk and inhibits β-catenin/TCF signaling. (C) Frizzled receptors in planar cell polarity (PCP) of vertebrates act via downstream messengers including Dishevelled (Dvl), small Rho GTPases, Jnk, or Ca2+. In flies, PCP can be activated independently of Wnt-Frizzled binding (?). (D) Binding of Wnt proteins to RYK RTKs results in the activation of Src proteins. (Adapted from van Amerongen and Nusse 2009; reprinted, with permission, from the authors.)
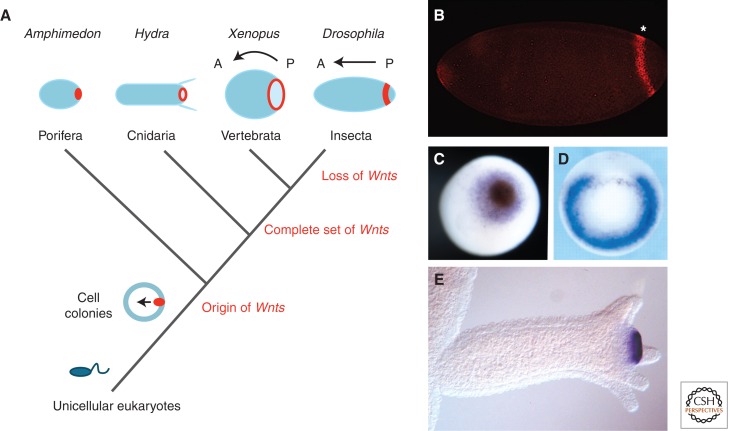
Evolution of Wnt signaling and axis formation during metazoan evolution. (A) Wnt signaling centers (red) evolved at the transition from unicellular (e.g., Monosiga) to multicellular eukaryotes. In all studied metazoans (sponges, cnidarians, deuterostomes, and protostomes), a posterior Wnt signaling center defines the posterior pole of the body axis. The number of Wnt gene subfamilies increased from the first metazoans to cnidarians, whose complete set of Wnt genes is only retained in basal deuterostomes and protostomes. Protostomes are characterized by frequent loss of Wnt gene subfamilies. (B) The posterior Wg (Wnt1) stripe in Drosophila. (Figure kindly provided by Drs. P. Vorwald and E. De Robertis, Howard Hughes Medical Institute, UC Los Angeles.) (C) Wnt expression at the blastopore of a Amphimedon gastrulae (AmqWntA), at the blastopore of an early Xenopus gastrulae (Wnt8) (figure kindly provided by Dr. Maja Adamska, SARS Center, Bergen), and (D) and (E) at the hypostome of Hydra polyps (Wnt3). (Fig. 2D from Steiner et al. 2006; reprinted, with permission, from The Company of Biologists © 2006; Fig. 2E from Hobmayer et al. 2000; reprinted, with permission, from the author.)
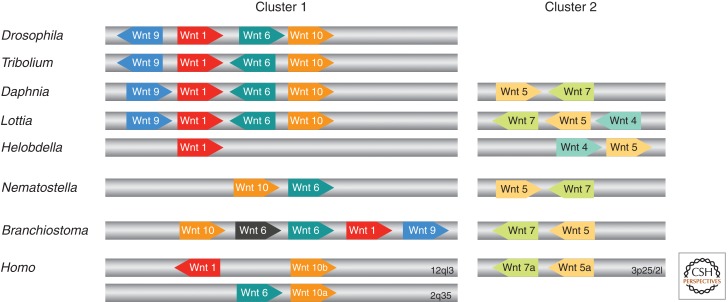
Synteny of metazoan Wnt genes. The relative position and orientation of Wnt genes with common scaffolds/chromosomes are shown for D. melanogaster, Tribolium castaneum, D. pulex, L. gigantea, H. robusta, Nematostella vectensis, Branchiostoma floridae, and Homo sapiens (not to scale). Numbers indicate specific chromosomes in the case of H. sapiens. (Figure created from data from Nusse [2001], Sullivan et al. [2007], Cho et al. [2010], and Janssen et al. [2010].) Note that there are more Wnts for a given species that are not clustered (see Table 1).
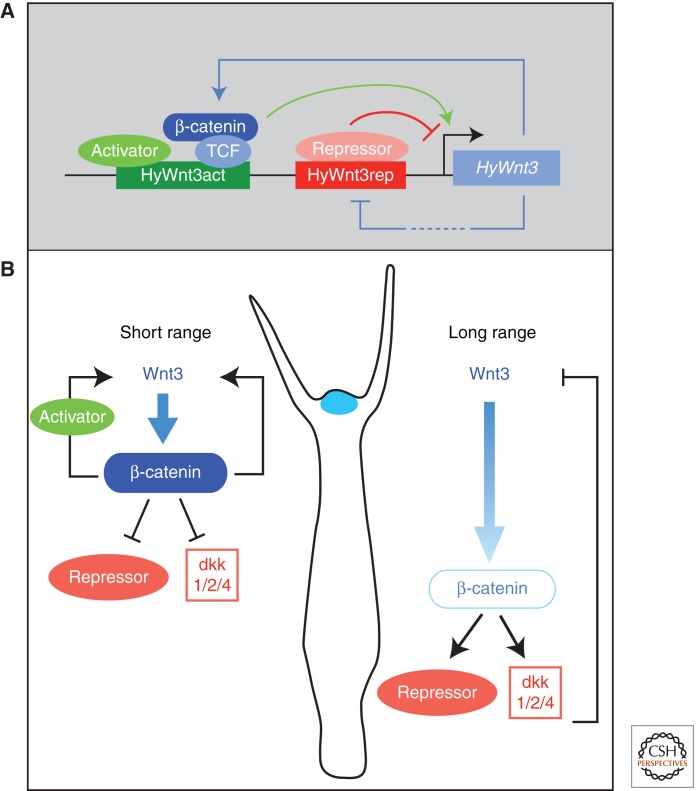
Transcriptional control of Wnt signaling in Hydra. (A) Model showing the transcriptional regulation of head organizer-specific HyWnt3 expression. HyWnt3 expression is controlled by two distinct cis-regulatory elements; the activator (HyWnt3act, green) and the repressor (HyWnt3rep, red) are positively and negatively regulated by Wnt/β-catenin signaling, respectively. The β-catenin/TCF complex and putative activators (light green) bind to HyWnt3act, and their combinatorial inputs act in HyWnt3 transcription (green arrow). Potential repressors (red) bind to a repressor element (red) and inhibit HyWnt3 expression. (Figure adapted from Nakamura et al. 2011; reprinted, with permission, from the author.) (B) Presumed distribution or activity of β-catenin depends on the diffusion range of Wnt3. Following short-range diffusion, the concentrations of Wnt3 and β-catenin are high, thereby repressing putative repressors and dkk1/2/4. Following long-range diffusion, the level of nuclear β-catenin is lower, inducing putative repressors including dkk1/2/4. Positive and negative regulation restricts HyWnt3 expression (blue) to the head organizer region.
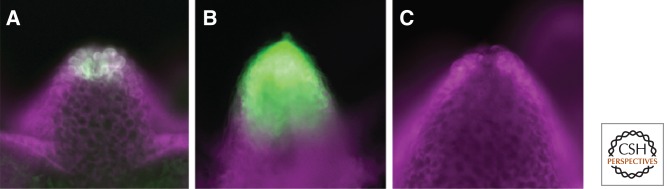
Expression of Wnt3 in the Hydra organizer. Reporter constructs for HyWnt3-EGFP (green) were used with an independent transformation marker (HyActin–RFP reporter gene, magenta) to ensure that all cells carry the EGFP reporter gene under examination. (A) The transgenic Hydra strain shows the localization of Wnt3 in epithelial cells at the tip of the hypostome with a complete Wnt3 promoter (HyWnt3FL). (B) Transgenic Hydra with a reporter construct lacking the HyWnt3 repressor element show dramatic expansion of expression. (C) Transgenic Hydra with a reporter construct lacking the HyWnt3 activator sequence including TCF binding sites show no expression. (From Nakamura et al. 2011; reprinted, with permission, from the author.)
References
-
- Abedin M, King N 2008. The premetazoan ancestry of cadherins. Science 319: 946–948 - PubMed
-
- Adamska M, Larroux C, Adamski M, Green K, Lovas E, Koop D, Richards GS, Zwafink C, Degnan BM 2010. Structure and expression of conserved Wnt pathway components in the demosponge Amphimedon queenslandica. Evol Dev 12: 494–518 - PubMed
-
- Agathon A, Thisse C, Thisse B 2003. The molecular nature of the zebrafish tail organizer. Nature 424: 448–452 - PubMed
-
- Arendt D, Technau U, Wittbrodt J 2001. Evolution of the bilaterian larval foregut. Nature 409: 81–85 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources