

Palmitoylation regulates vesicular trafficking of R-Ras to membrane ruffles and effects on ruffling and cell spreading
- PMID: 22751447
- PMCID: PMC3442799
- DOI: 10.4161/sgtp.21084
Palmitoylation regulates vesicular trafficking of R-Ras to membrane ruffles and effects on ruffling and cell spreading
Abstract
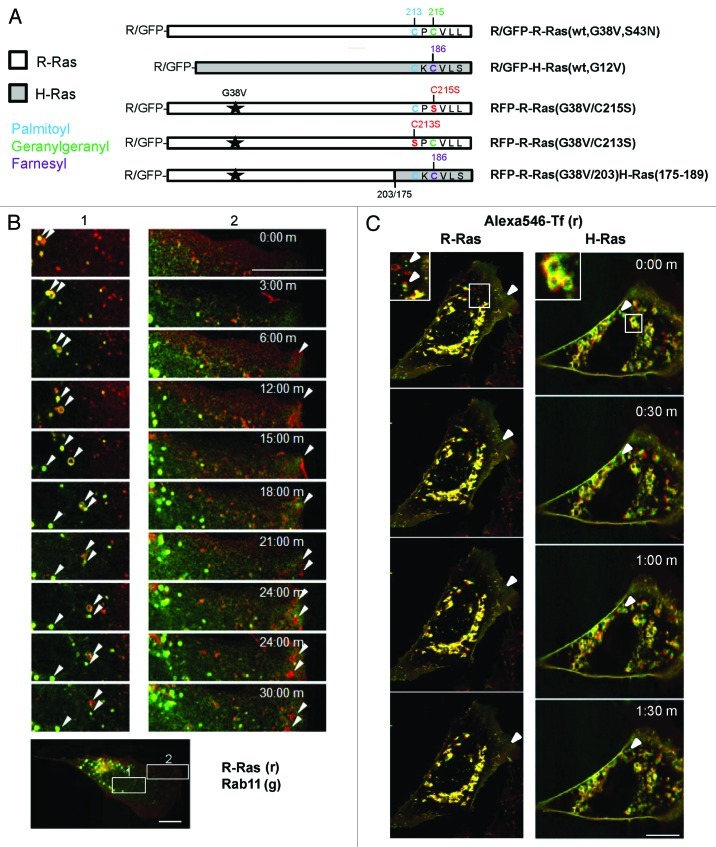
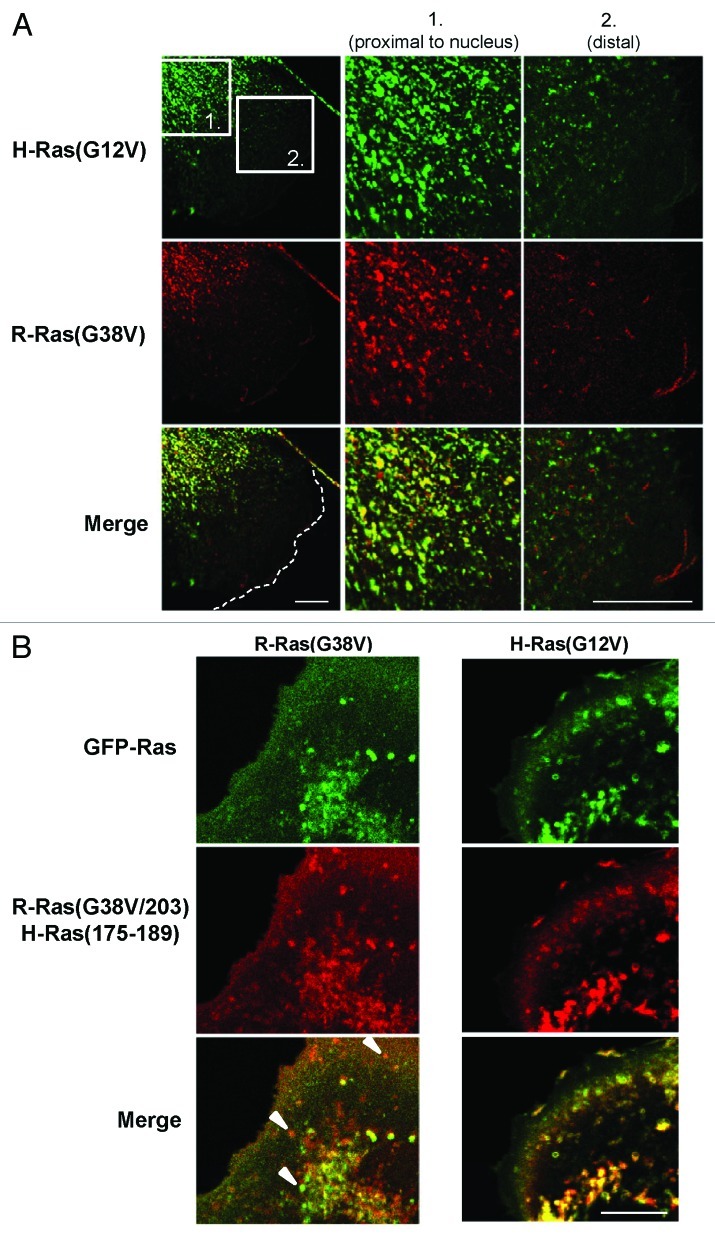
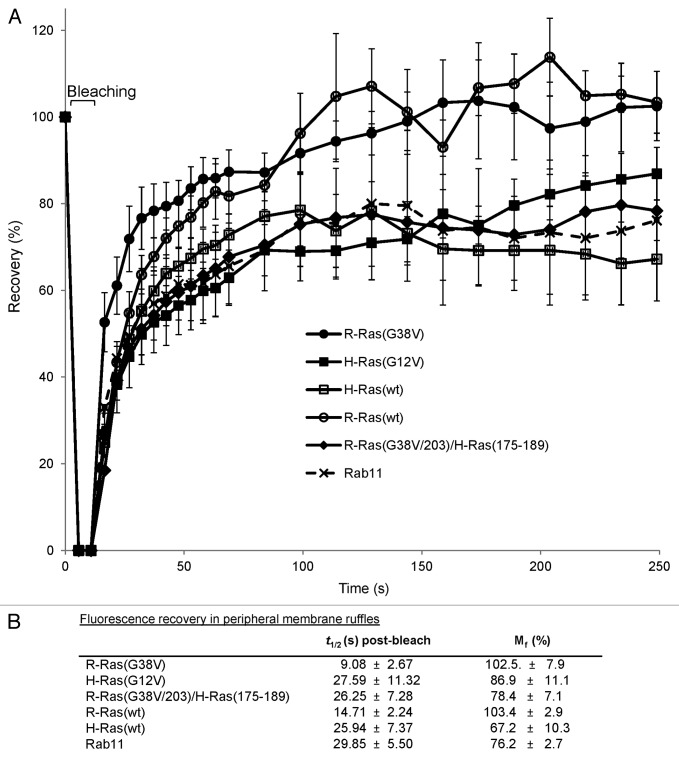
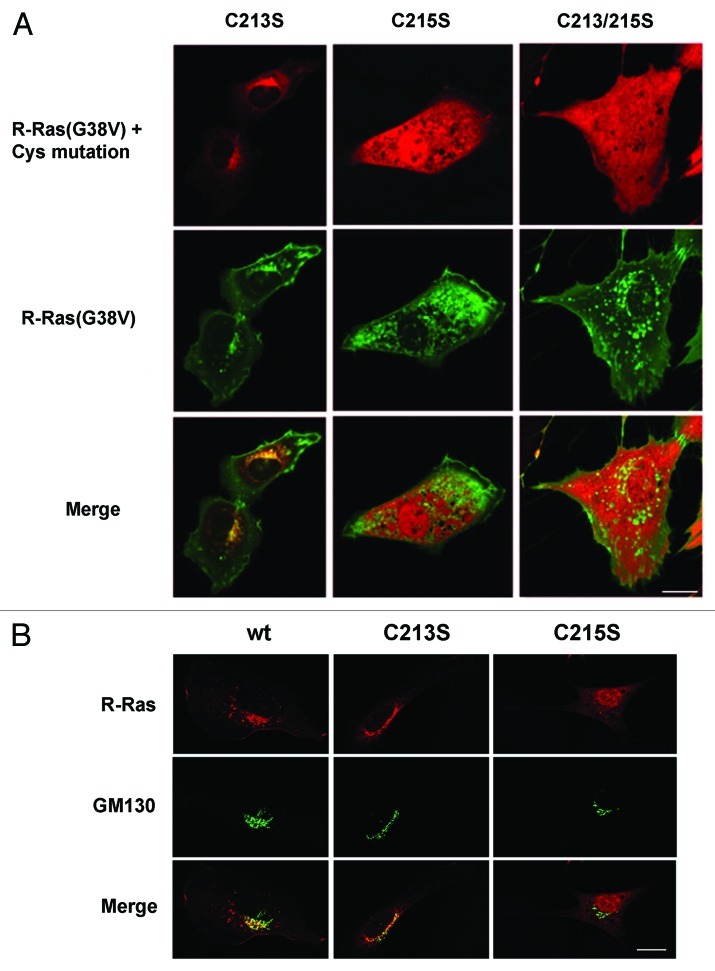
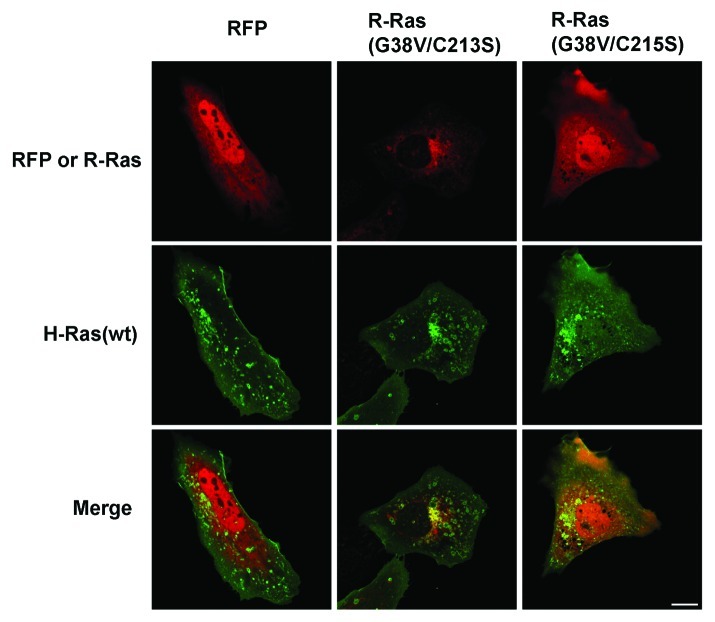
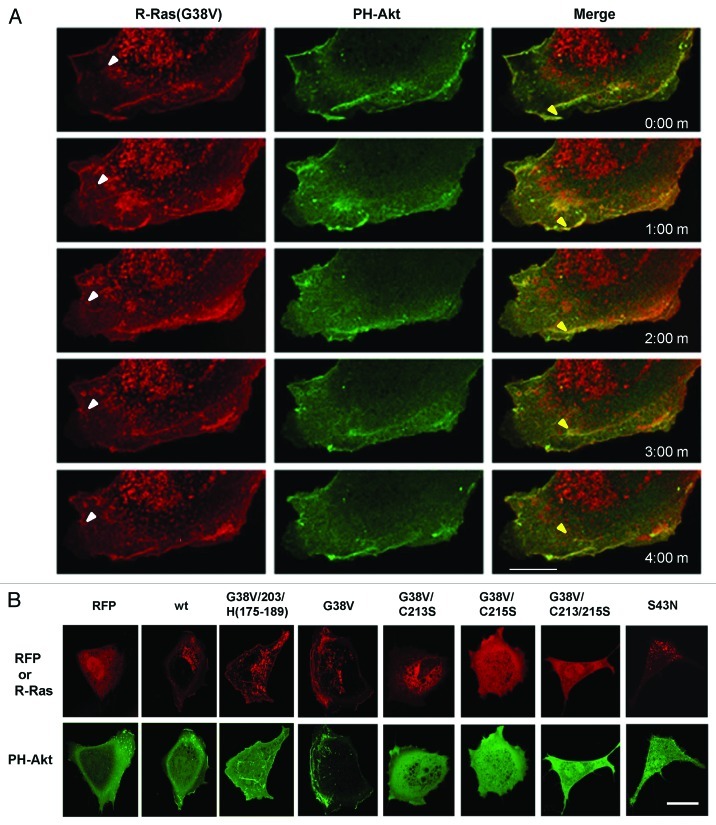
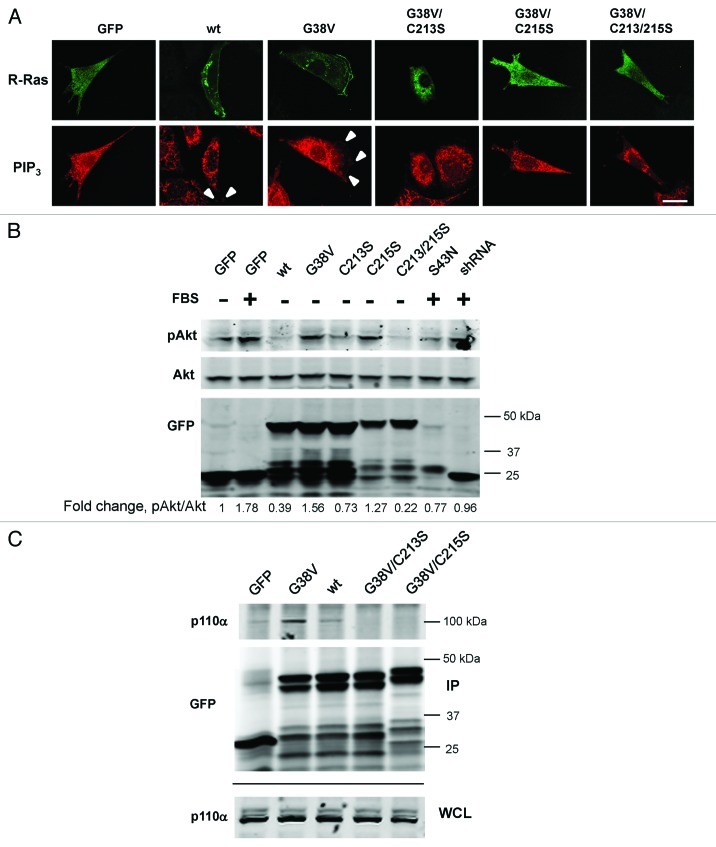
In this study we investigated the dynamics of R-Ras intracellular trafficking and its contributions to the unique roles of R-Ras in membrane ruffling and cell spreading. Wild type and constitutively active R-Ras localized to membranes of both Rab11- and transferrin-positive and -negative vesicles, which trafficked anterograde to the leading edge in migrating cells. H-Ras also co-localized with R-Ras in many of these vesicles in the vicinity of the Golgi, but R-Ras and H-Ras vesicles segregated proximal to the leading edge, in a manner dictated by the C-terminal membrane-targeting sequences. These segregated vesicle trafficking patterns corresponded to distinct modes of targeting to membrane ruffles at the leading edge. Geranylgeranylation was required for membrane anchorage of R-Ras, whereas palmitoylation was required for exit from the Golgi in post-Golgi vesicle membranes and trafficking to the plasma membrane. R-Ras vesicle membranes did not contain phosphatidylinositol (3,4,5)-trisphosphate (PtdIns(3,4,5)P(3)), whereas R-Ras co-localized with PtdIns(3,4,5)P(3) in membrane ruffles. Finally, palmitoylation-deficient R-Ras blocked membrane ruffling, R-Ras/PI3-kinase interaction, enrichment of PtdIns(3,4,5)P(3) at the plasma membrane, and R-Ras-dependent cell spreading. Thus, lipid modification of R-Ras dictates its vesicle trafficking, targeting to membrane ruffles, and its unique roles in localizing PtdIns(3,4,5)P(3) to ruffles and promoting cell spreading.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous