

For many but not for all: how the conformational flexibility of the peptide/MHCII complex shapes epitope selection
- PMID: 22753017
- PMCID: PMC4197051
- DOI: 10.1007/s12026-012-8342-2
For many but not for all: how the conformational flexibility of the peptide/MHCII complex shapes epitope selection
Abstract
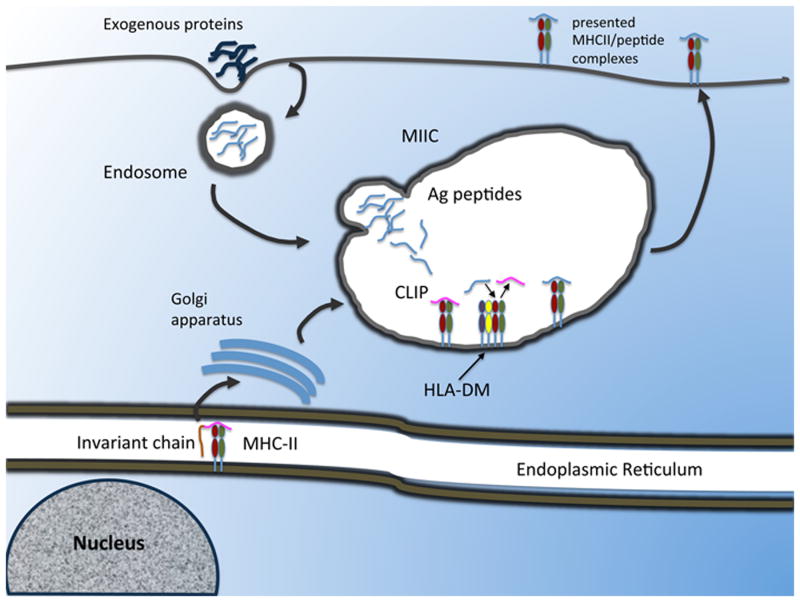
The adaptive immune response starts when CD4+ T cells recognize peptide antigens presented by class II molecules of the Major Histocompatibility Complex (MHCII). Two outstanding features of MHCII molecules are their polymorphism and the ability of each allele to bind a large panoply of peptides. The ability of each MHCII molecule to interact with a limited, though broad, range of amino acid sequences, or "permissive specificity" of binding, is the result of structural flexibility. This flexibility has been identified through biochemical and biophysical studies, and molecular dynamic simulations have modeled the conformational rearrangements that the peptide and the MHCII undergo during interaction. Moreover, there is evidence that the structural flexibility of the peptide/MHCII complex correlates with the activity of the "peptide-editing" molecule DM. In light of the impact that these recent findings have on our ability to predict MHCII epitopes, a review of the structural and thermodynamic determinants of peptide binding to MHCII is proposed.
Figures
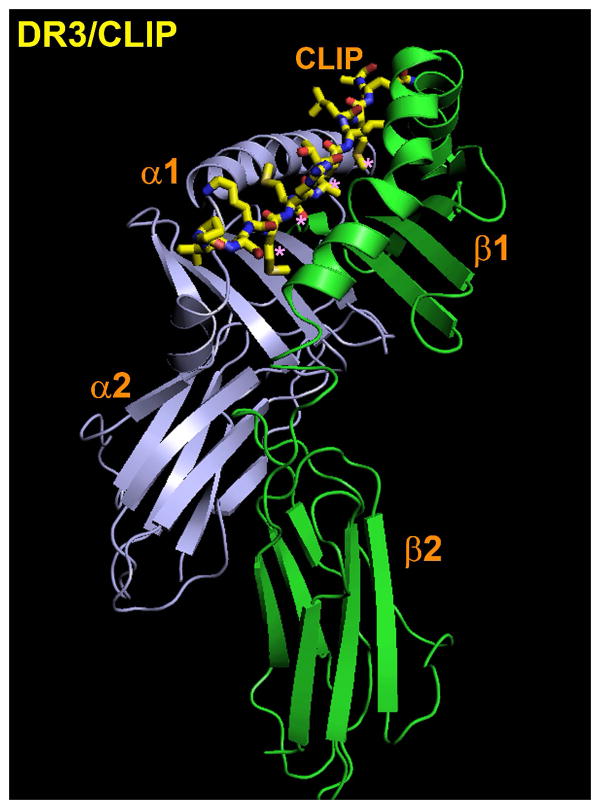
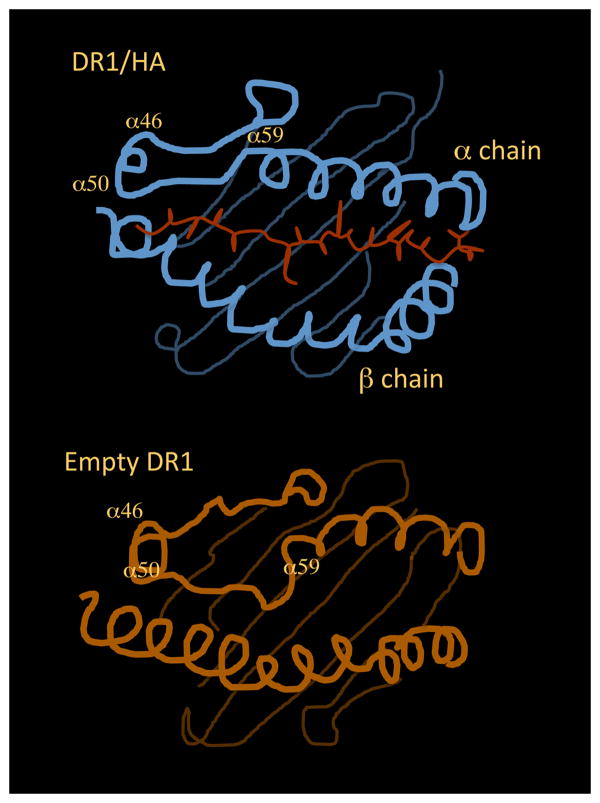
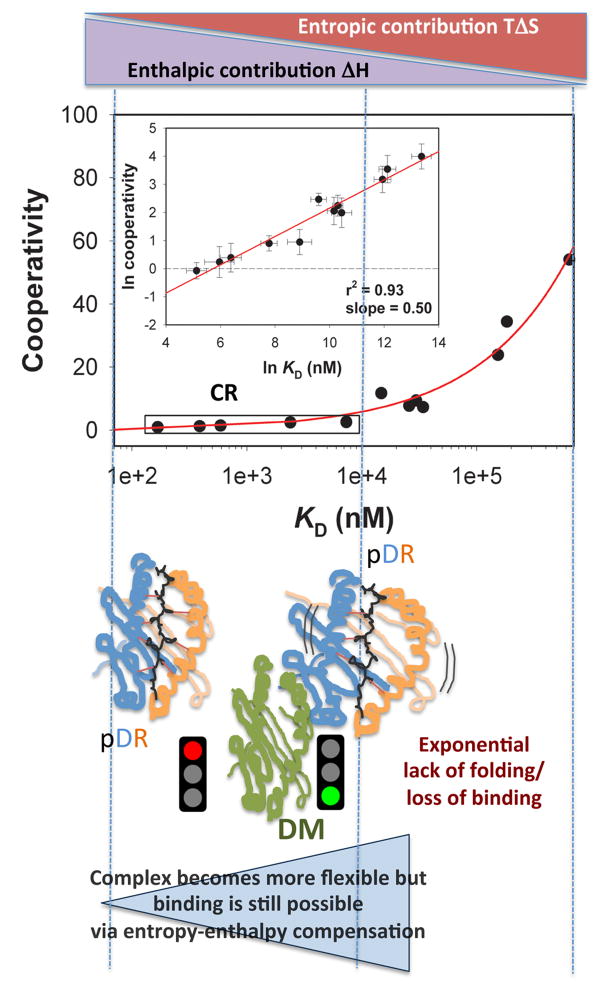
 . The impact of cooperative effects on peptide affinity has been investigated applying the mutant cycle approach to the DR/HA system. Due tothe disruptive nature of the modifications, the observed Cooperativity can be interpreted as lack of folding. Cooperativity affects complex formation in an exponential fashion, indicating that disrupting interaction after interaction has an amplified effect on the ability of the complex to fold into a stable conformer. At the left side of the curve, an affinity range can be identified, for which null or little cooperativity can be measured (Compensatory Range, CR framed in the plot). The broader is this range, the greater is the ability of the system to compensate any lack of interactions (H-bonds, Hydrophobic or salt bridges) with the residual flexibility (phenomenon of entropy-enthalpy compensation). To the extent that DM interacts and destabilizes complexes featuring greater residual entropy, thus reducing the compensatory range for stable complexion, this approach may also be used to identify the susceptibility to DM of a complex on the basis of its thermodynamic profile.
. The impact of cooperative effects on peptide affinity has been investigated applying the mutant cycle approach to the DR/HA system. Due tothe disruptive nature of the modifications, the observed Cooperativity can be interpreted as lack of folding. Cooperativity affects complex formation in an exponential fashion, indicating that disrupting interaction after interaction has an amplified effect on the ability of the complex to fold into a stable conformer. At the left side of the curve, an affinity range can be identified, for which null or little cooperativity can be measured (Compensatory Range, CR framed in the plot). The broader is this range, the greater is the ability of the system to compensate any lack of interactions (H-bonds, Hydrophobic or salt bridges) with the residual flexibility (phenomenon of entropy-enthalpy compensation). To the extent that DM interacts and destabilizes complexes featuring greater residual entropy, thus reducing the compensatory range for stable complexion, this approach may also be used to identify the susceptibility to DM of a complex on the basis of its thermodynamic profile.References
-
- Benacerraf B. Role of MHC gene products in immune regulation. Science. 1981;212(4500):1229–38. Epub 1981/06/12. - PubMed
-
- Nelson CA, Fremont DH. Structural principles of MHC class II antigen presentation. Rev Immunogenet. 1999;1(1):47–59. - PubMed
-
- Chaturvedi P, Yu Q, Southwood S, Sette A, Singh B. Peptide analogs with different affinites for MHC alter the cytokine profile of T helper cells. Int Immunol. 1996;8(5):745–55. - PubMed
-
- Engelhard VH. Structure of peptides associated with class I and class II MHC molecules. Annu Rev Immunol. 1994;12:181–207. - PubMed
-
- Lippolis JD, White FM, Marto JA, Luckey CJ, Bullock TN, Shabanowitz J, et al. Analysis of MHC class II antigen processing by quantitation of peptides that constitute nested sets. J Immunol. 2002;169(9):5089–97. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
