

Hypoxic regulation of cardiac Ca2+ channel: possible role of haem oxygenase
- PMID: 22753548
- PMCID: PMC3473281
- DOI: 10.1113/jphysiol.2012.236570
Hypoxic regulation of cardiac Ca2+ channel: possible role of haem oxygenase
Abstract
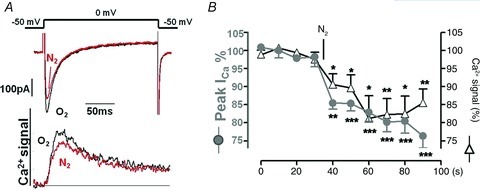
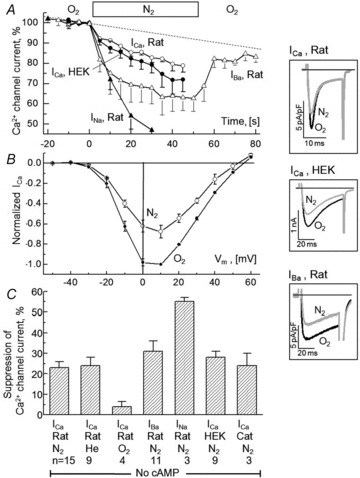
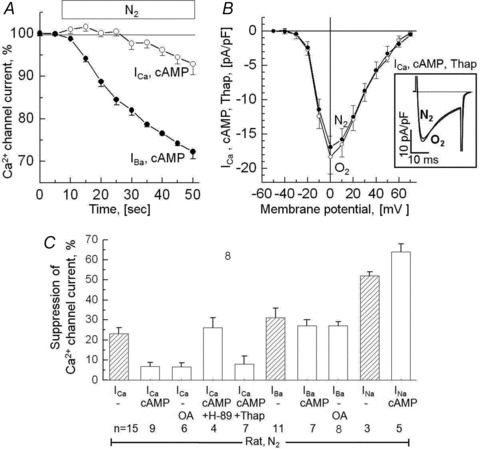
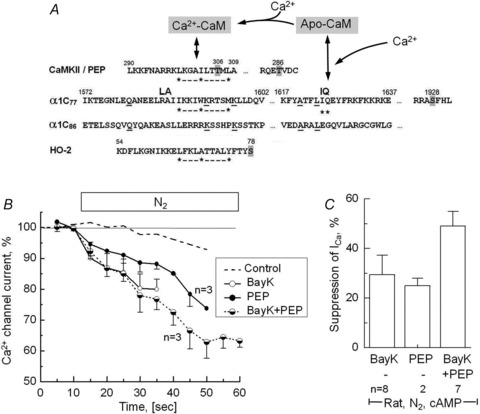
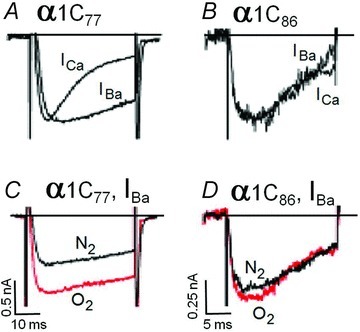
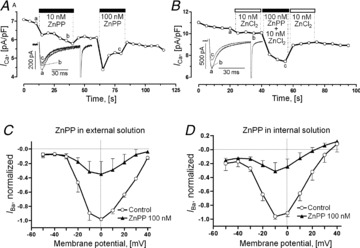
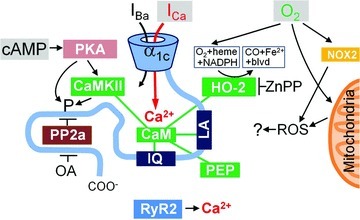
Acute and chronic hypoxias are common cardiac diseases that lead often to arrhythmia and impaired contractility. At the cellular level it is unclear whether the suppression of cardiac Ca(2+) channels (Ca(V)1.2) results directly from oxygen deprivation on the channel protein or is mediated by intermediary proteins affecting the channel. To address this question we measured the early effects of hypoxia (5-60 s, P(O(2)) < 5 mmHg) on Ca(2+) current (I(Ca)) and tested the involvement of protein kinase A (PKA) phosphorylation, Ca(2+)/calmodulin-mediated signalling and the haem oxygenase (HO) pathway in the hypoxic regulation of Ca(V)1.2 in rat and cat ventricular myocytes and HEK-293 cells. Hypoxic suppression of ICa) and Ca(2+) transients was significant within 5 s and intensified in the following 50 s, and was reversible. Phosphorylation by cAMP or the phosphatase inhibitor okadaic acid desensitized I(Ca) to hypoxia, while PKA inhibition by H-89 restored the sensitivity of I(Ca) to hypoxia. This phosphorylation effect was specific to Ca(2+), but not Ba(2+) or Na(+), permeating through the channel. CaMKII inhibitory peptide and Bay K8644 reversed the phosphorylation-induced desensitization to hypoxia. Mutation of CAM/CaMKII-binding motifs of the α(1c) subunit of Ca(V)1.2 fully desensitized the Ca(2+) channel to hypoxia. Rapid application of HO inhibitors (zinc protoporphyrin (ZnPP) and tin protoporphyrin (SnPP)) suppressed the channel in a manner similar to acute hypoxia such that: (1) I(Ca) and I(Ba) were suppressed within 5 s of ZnPP application; (2) PKA activation and CaMKII inhibitors desensitized I(Ca), but not I(Ba), to ZnPP; and (3) hypoxia failed to further suppress I(Ca) and I(Ba) in ZnPP-treated myocytes. We propose that the binding of HO to the CaM/CaMKII-specific motifs on Ca(2+) channel may mediate the rapid response of the channel to hypoxia.
Figures

References
-
- Adachi-Akahane S, Cleemann L, Morad M. BAY K 8644 modifies Ca2+ cross signaling between DHP and ryanodine receptors in rat ventricular myocytes. Am J Physiol Heart Circ Physiol. 1999;276:H1178–1189. - PubMed
-
- Belmonte S, Morad M. Shear fluid-induced Ca2+ release and the role of mitochondria in rat cardiac myocytes. Ann NY Acad Sci. 2008;1123:58–63. - PubMed
-
- Boehning D, Sedaghat L, Sedlak TW, Snyder SH. Heme oxygenase-2 is activated by calcium-calmodulin. J Biol Chem. 2004;279:30927–30930. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous