

Constitutive activity of TRPML2 and TRPML3 channels versus activation by low extracellular sodium and small molecules
- PMID: 22753890
- PMCID: PMC3391124
- DOI: 10.1074/jbc.M112.368876
Constitutive activity of TRPML2 and TRPML3 channels versus activation by low extracellular sodium and small molecules
Abstract
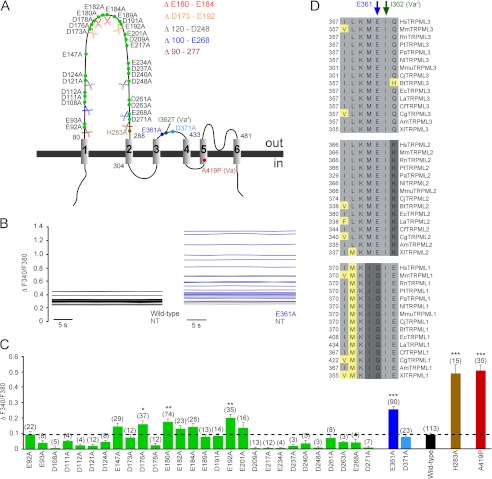
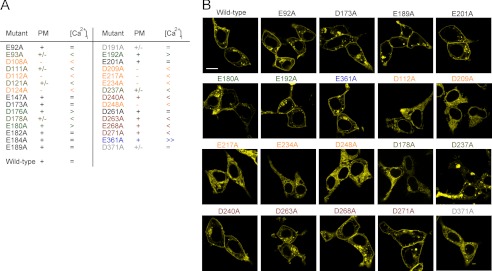
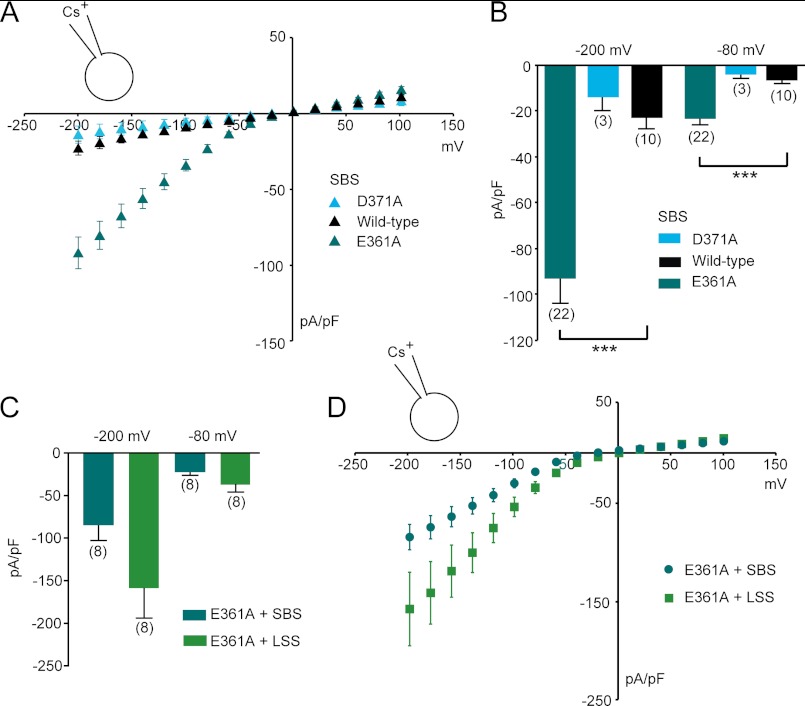
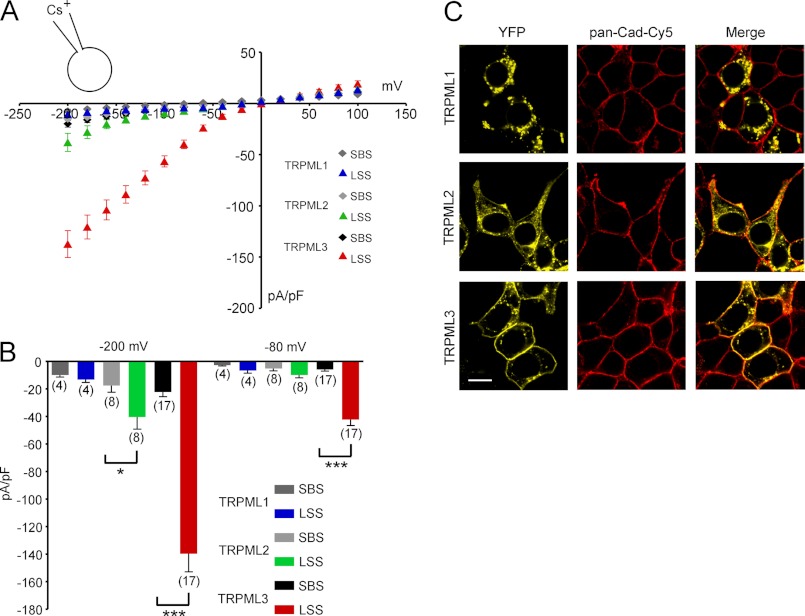
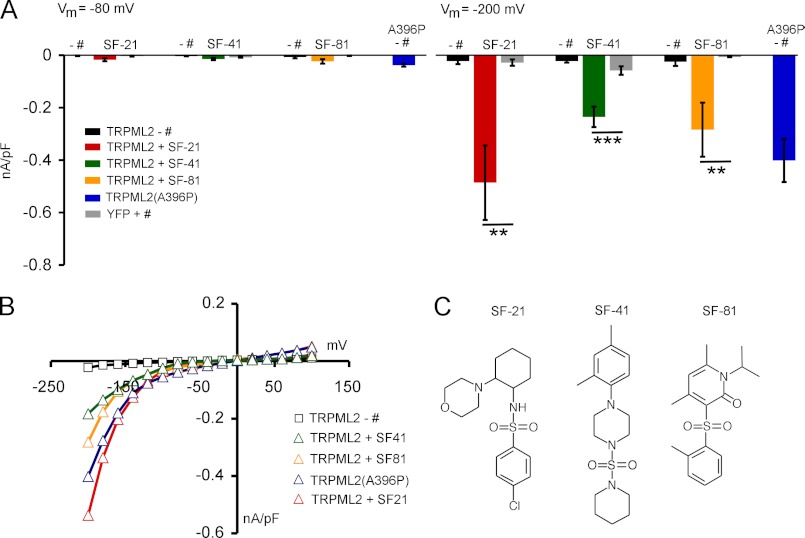
The transient receptor potential channels TRPML2 and TRPML3 (MCOLN2 and MCOLN3) are nonselective cation channels. They are widely expressed in mammals. However, little is known about their physiological function(s) and activation mechanism(s). TRPML3 can be activated or rather de-inhibited by exposing it first to sodium-free extracellular solution and subsequently to high extracellular sodium. TRPML3 can also be activated by a variety of small chemical compounds identified in a high throughput screen and is inhibited by low pH. Furthermore, it was found that TRPML3 is constitutively active in low or no sodium-containing extracellular solution. This constitutive activity is independent of the intracellular presence of sodium, and whole-cell current densities are similar with pipette solutions containing cesium, potassium, or sodium. Here, we present mutagenesis data generated based on the hypothesis that negatively charged amino acids in the extracellular loops of TRPML3 may interfere with the observed sodium inhibition. We systematically mutated negatively charged amino acids in the first and second extracellular loops and found that mutating Glu-361 in the second loop has a significant impact on the sodium-mediated block of TRPML3. We further demonstrate that the TRPML3-related cation channel TRPML2 is also activated by lowering the extracellular sodium concentration as well as by a subset of small chemical compounds that were previously identified as activators of TRPML3, thus confirming the functional activity of TRPML2 at the plasma membrane and suggesting similar gating mechanisms for both TRPML channels.
Figures
Similar articles
-
TRPML2 and mucolipin evolution.Handb Exp Pharmacol. 2014;222:647-58. doi: 10.1007/978-3-642-54215-2_25. Handb Exp Pharmacol. 2014. PMID: 24756724 Review.
-
Constitutive activity of the human TRPML2 channel induces cell degeneration.J Biol Chem. 2010 Jan 22;285(4):2771-82. doi: 10.1074/jbc.M109.046508. Epub 2009 Nov 23. J Biol Chem. 2010. PMID: 19940139 Free PMC article.
-
Lysosomal localization of TRPML3 depends on TRPML2 and the mucolipidosis-associated protein TRPML1.J Biol Chem. 2006 Jun 23;281(25):17517-17527. doi: 10.1074/jbc.M600807200. Epub 2006 Apr 10. J Biol Chem. 2006. PMID: 16606612 Free PMC article.
-
The mucolipin-2 (TRPML2) ion channel: a tissue-specific protein crucial to normal cell function.Pflugers Arch. 2016 Feb;468(2):177-92. doi: 10.1007/s00424-015-1732-2. Epub 2015 Sep 4. Pflugers Arch. 2016. PMID: 26336837 Free PMC article. Review.
-
Palmitoylation controls trafficking of the intracellular Ca2+ channel MCOLN3/TRPML3 to regulate autophagy.Autophagy. 2019 Feb;15(2):327-340. doi: 10.1080/15548627.2018.1518671. Epub 2018 Sep 14. Autophagy. 2019. PMID: 30215288 Free PMC article.
Cited by
-
Cryo-EM structures of the human endolysosomal TRPML3 channel in three distinct states.Nat Struct Mol Biol. 2017 Dec;24(12):1146-1154. doi: 10.1038/nsmb.3502. Epub 2017 Nov 6. Nat Struct Mol Biol. 2017. PMID: 29106414 Free PMC article.
-
Estradiol analogs attenuate autophagy, cell migration and invasion by direct and selective inhibition of TRPML1, independent of estrogen receptors.Sci Rep. 2021 Apr 15;11(1):8313. doi: 10.1038/s41598-021-87817-4. Sci Rep. 2021. PMID: 33859333 Free PMC article.
-
The Dictyostelium Model for Mucolipidosis Type IV.Front Cell Dev Biol. 2022 Apr 13;10:741967. doi: 10.3389/fcell.2022.741967. eCollection 2022. Front Cell Dev Biol. 2022. PMID: 35493081 Free PMC article.
-
Polycystin-2 is an essential ion channel subunit in the primary cilium of the renal collecting duct epithelium.Elife. 2018 Feb 14;7:e33183. doi: 10.7554/eLife.33183. Elife. 2018. PMID: 29443690 Free PMC article.
-
Selective agonist of TRPML2 reveals direct role in chemokine release from innate immune cells.Elife. 2018 Nov 27;7:e39720. doi: 10.7554/eLife.39720. Elife. 2018. PMID: 30479274 Free PMC article.
References
-
- van Aken A. F., Atiba-Davies M., Marcotti W., Goodyear R. J., Bryant J. E., Richardson G. P., Noben-Trauth K., Kros C. J. (2008) TRPML3 mutations cause impaired mechano-electrical transduction and depolarization by an inward-rectifier cation current in auditory hair cells of varitint-waddler mice. J. Physiol. 586, 5403–5418 - PMC - PubMed
-
- Takumida M., Anniko M. (2010) Expression of transient receptor potential channel mucolipin (TRPML) and polycystine (TRPP) in the mouse inner ear. Acta Otolaryngol. 130, 196–203 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
