

Interactions between epinephrine, ascending vagal fibers, and central noradrenergic systems in modulating memory for emotionally arousing events
- PMID: 22754515
- PMCID: PMC3384987
- DOI: 10.3389/fnbeh.2012.00035
Interactions between epinephrine, ascending vagal fibers, and central noradrenergic systems in modulating memory for emotionally arousing events
Abstract
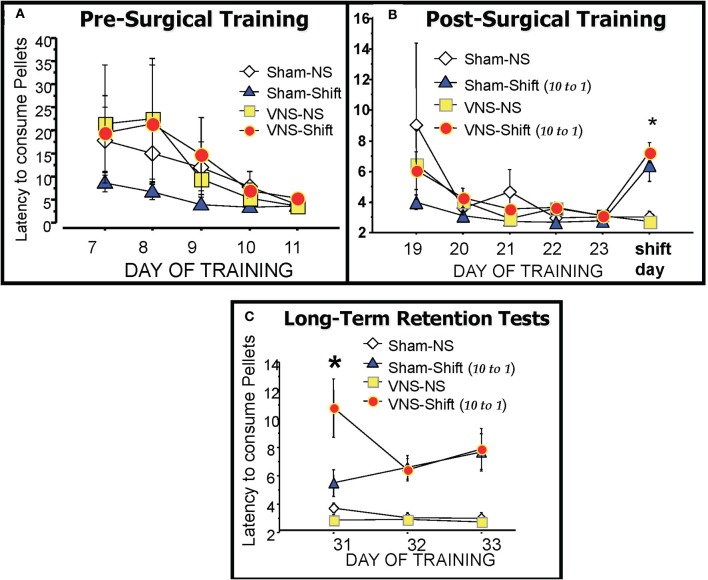
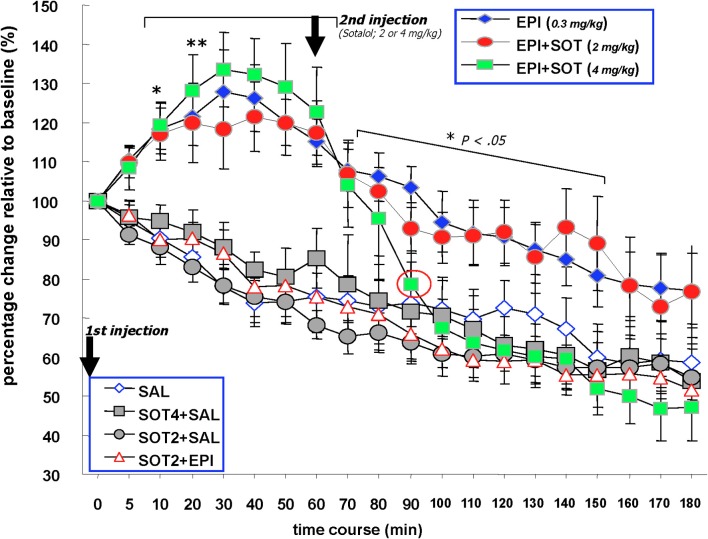
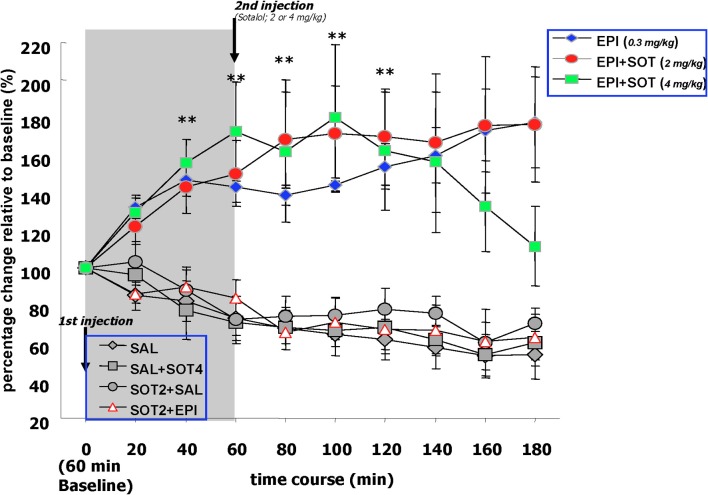
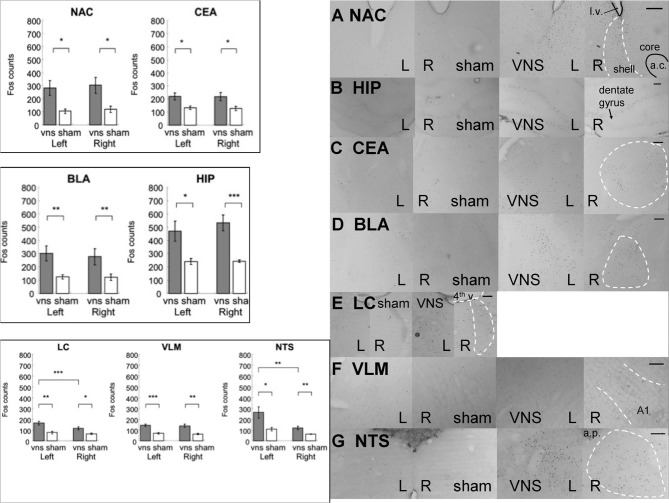
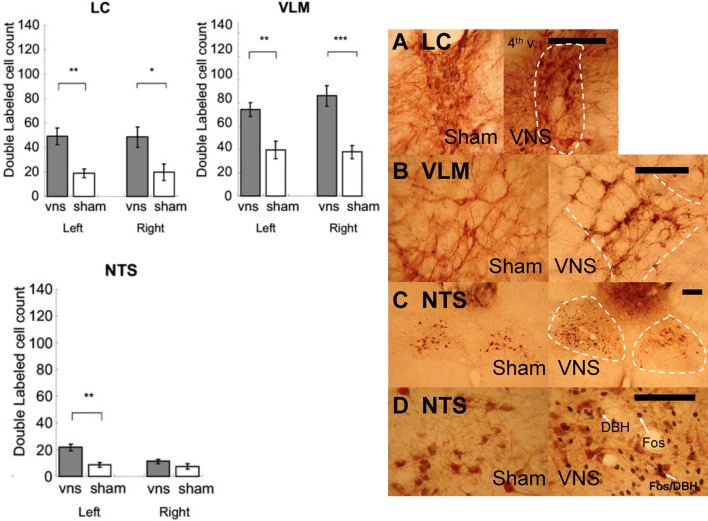
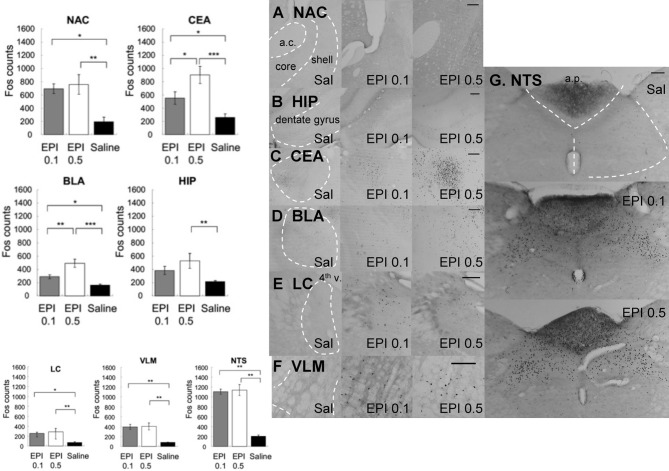
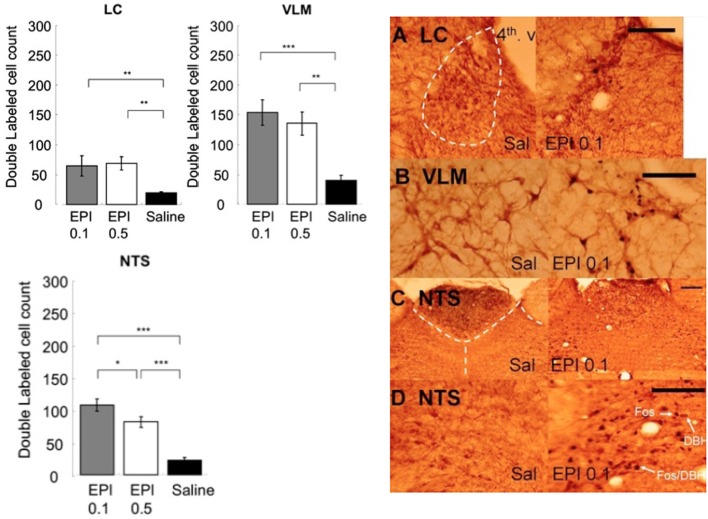
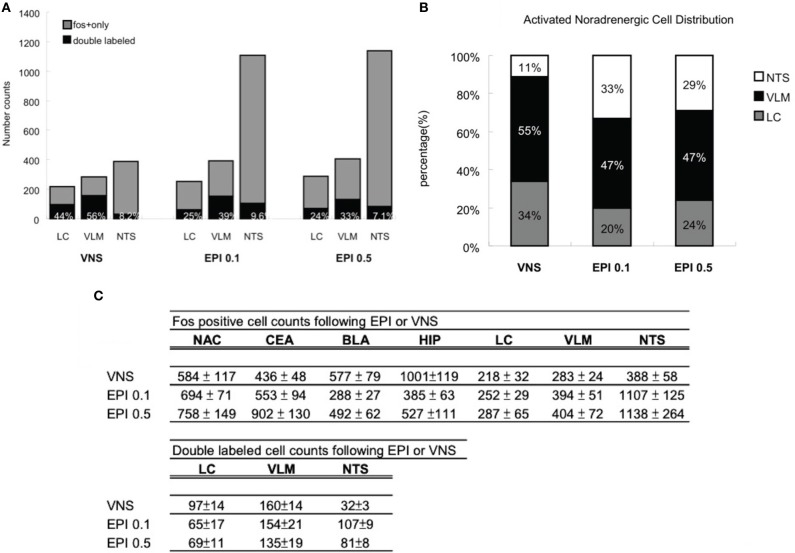
It is well-established that exposure to emotionally laden events initiates secretion of the arousal-related hormone epinephrine in the periphery. These neuroendocrine changes and the subsequent increase in peripheral physiological output play an integral role in modulating brain systems involved in memory formation. The impermeability of the blood brain barrier to epinephrine represents an important obstacle in understanding how peripheral hormones initiate neurochemical changes in the brain that lead to effective memory formation. This obstacle necessitated the identity of a putative pathway capable of conveying physiological changes produced by epinephrine to limbic structures that incorporate arousal and affect related information into memory. A major theme of the proposed studies is that ascending fibers of the vagus nerve may represent such a mechanism. This hypothesis was tested by evaluating the contribution of ascending vagal fibers in modulating memory for responses learned under behavioral conditions that produce emotional arousal by manipulating appetitive stimuli. A combination of electrophysiological recording of vagal afferent fibers and in vivo microdialysis was employed in a second study to simultaneously assess how elevations in peripheral levels of epinephrine affect vagal nerve discharge and the subsequent potentiation of norepinephrine release in the basolateral amygdala. The final study used double immunohistochemistry labeling of c-fos and dopamine beta hydroxylase (DBH), the enzyme for norepinephrine synthesis to determine if epinephrine administration alone or stimulation of the vagus nerve at an intensity identical to that which improved memory in Experiment 1 produces similar patterns of neuronal activity in brain areas involved in processing memory for emotional events. Findings emerging from this collection of studies establish the importance of ascending fibers of the vagus nerve as an essential pathway for conveying the peripheral consequences of physiological arousal on brain systems that encode new information into memory storage.
Keywords: amygdala; emotional arousal; epinephrine; learning; memory; vagus nerve.
Figures

Similar articles
-
Epinephrine administration increases neural impulses propagated along the vagus nerve: Role of peripheral beta-adrenergic receptors.Neurobiol Learn Mem. 2006 Mar;85(2):116-24. doi: 10.1016/j.nlm.2005.08.013. Epub 2005 Oct 17. Neurobiol Learn Mem. 2006. PMID: 16230035
-
The effects of peripheral vagal nerve stimulation at a memory-modulating intensity on norepinephrine output in the basolateral amygdala.Behav Neurosci. 2004 Feb;118(1):79-88. doi: 10.1037/0735-7044.118.1.79. Behav Neurosci. 2004. PMID: 14979784
-
Role of amygdala norepinephrine in mediating stress hormone regulation of memory storage.Acta Pharmacol Sin. 2000 Jun;21(6):481-93. Acta Pharmacol Sin. 2000. PMID: 11360681 Review.
-
Role of the basolateral amygdala in memory consolidation.Ann N Y Acad Sci. 2003 Apr;985:273-93. doi: 10.1111/j.1749-6632.2003.tb07088.x. Ann N Y Acad Sci. 2003. PMID: 12724165 Review.
-
Emotionally arousing pictures increase blood glucose levels and enhance recall.Neurobiol Learn Mem. 2001 May;75(3):262-73. doi: 10.1006/nlme.2000.3973. Neurobiol Learn Mem. 2001. PMID: 11300733
Cited by
-
Vagus nerve stimulation: a physical therapy with promising potential for central nervous system disorders.Front Neurol. 2024 Dec 13;15:1516242. doi: 10.3389/fneur.2024.1516242. eCollection 2024. Front Neurol. 2024. PMID: 39734634 Free PMC article. Review.
-
Stress and the "extended" autonomic system.Auton Neurosci. 2021 Dec;236:102889. doi: 10.1016/j.autneu.2021.102889. Epub 2021 Oct 2. Auton Neurosci. 2021. PMID: 34656967 Free PMC article. Review.
-
Neuromodulation and a Reconceptualization of Autism Spectrum Disorders: Using the Locus Coeruleus Functioning as an Exemplar.Front Neurol. 2018 Dec 19;9:1120. doi: 10.3389/fneur.2018.01120. eCollection 2018. Front Neurol. 2018. PMID: 30619071 Free PMC article.
-
Vagus nerve stimulation promotes extinction generalization across sensory modalities.Neurobiol Learn Mem. 2021 May;181:107425. doi: 10.1016/j.nlm.2021.107425. Epub 2021 Mar 23. Neurobiol Learn Mem. 2021. PMID: 33771710 Free PMC article.
-
Generation of Two Noradrenergic-Specific Dopamine-Beta-Hydroxylase-FLPo Knock-In Mice Using CRISPR/Cas9-Mediated Targeting in Embryonic Stem Cells.PLoS One. 2016 Jul 21;11(7):e0159474. doi: 10.1371/journal.pone.0159474. eCollection 2016. PLoS One. 2016. PMID: 27441631 Free PMC article.
References
-
- Benavides M., Laorden M. L., Milanes M. V. (2005). Involvement of 3′, 5′-cyclic adenosine monophosphate-dependent protein kinase in regulation of Fos expression and tyrosine hydroxylase levels during morphine withdrawal in the hypothalamic paraventricular nucleus and medulla oblongata catecholaminergic cell groups. J. Neurochem. 92, 246–254 10.1111/j.1471-4159.2004.02865.x - DOI - PubMed
-
- Bjorklund A., Lindvall O. (1986). “Catecholaminergic brain stem regulatory systems,” in Handbook of Physiology, A Critical, Comprehensive Presentation of Physiological Knowledge and Concepts, Section, I, The Nervous System, ed Mountcastle V. B. (Bethesda: American Physiological Society; ), 155–235
-
- Bohning D. E., Lomarev M. P., Denslow S., Nahas Z., Shastri A., George M. S. (2001). Feasibility of vagus nerve stimulation-synchronized blood oxygenation level-dependent functional MRI. Invest. Radiol. 36, 470–479 - PubMed
-
- Bradbury M. W. (1993). The blood-brain barrier. Exp. Physiol. 78, 453–472 - PubMed
LinkOut - more resources
Full Text Sources
Miscellaneous