

Horizontal gene transfer dynamics and distribution of fitness effects during microbial in silico evolution
- PMID: 22759418
- PMCID: PMC3382434
- DOI: 10.1186/1471-2105-13-S10-S13
Horizontal gene transfer dynamics and distribution of fitness effects during microbial in silico evolution
Abstract
Background: Horizontal gene transfer (HGT) is a process that facilitates the transfer of genetic material between organisms that are not directly related, and thus can affect both the rate of evolution and emergence of traits. Recent phylogenetic studies reveal HGT events are likely ubiquitous in the Tree of Life. However, our knowledge of HGT's role in evolution and biological organization is very limited, mainly due to the lack of ancestral evolutionary signatures and the difficulty to observe complex evolutionary dynamics in a laboratory setting. Here, we utilize a multi-scale microbial evolution model to comprehensively study the effect of HGT on the evolution of complex traits and organization of gene regulatory networks.
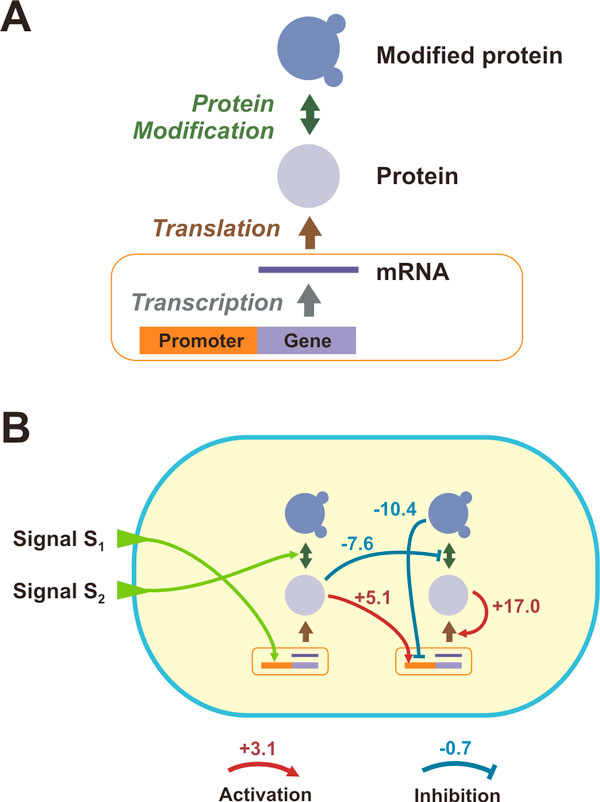
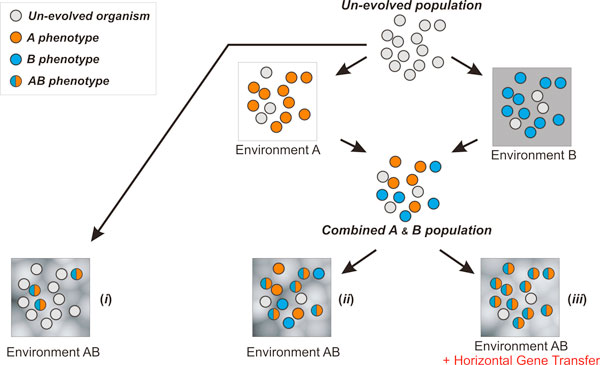
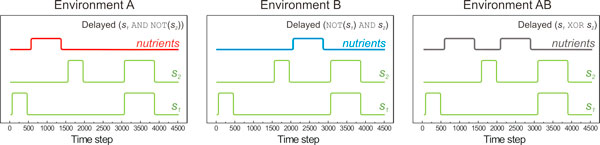
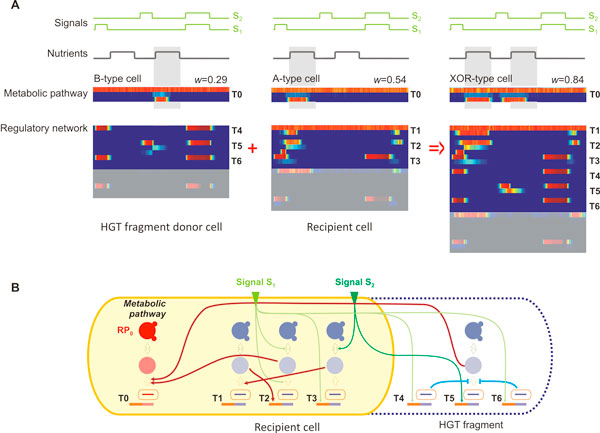
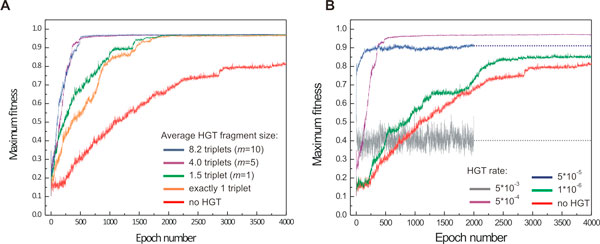
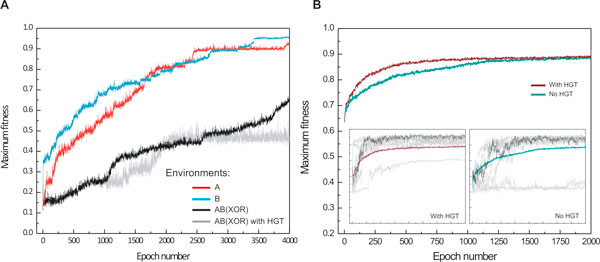
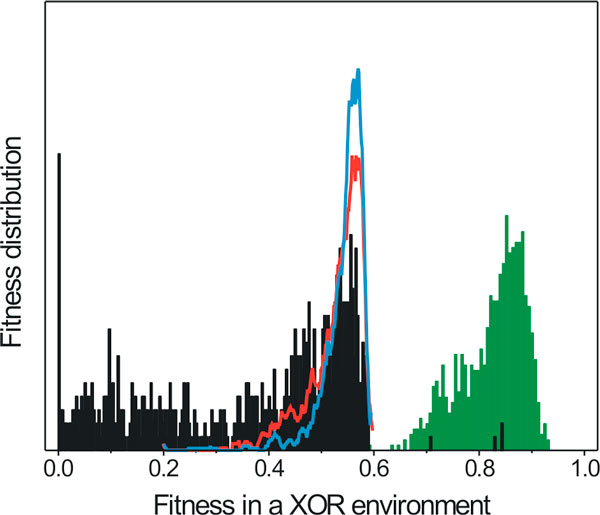
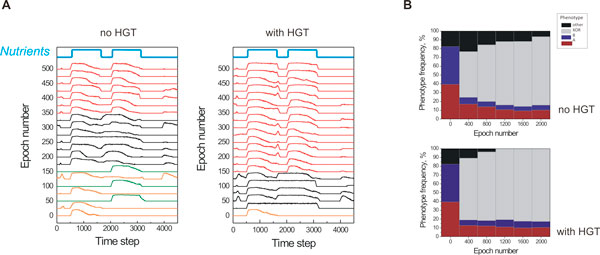
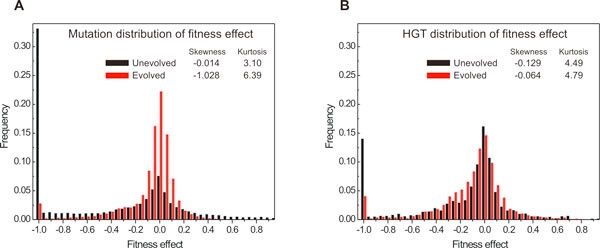
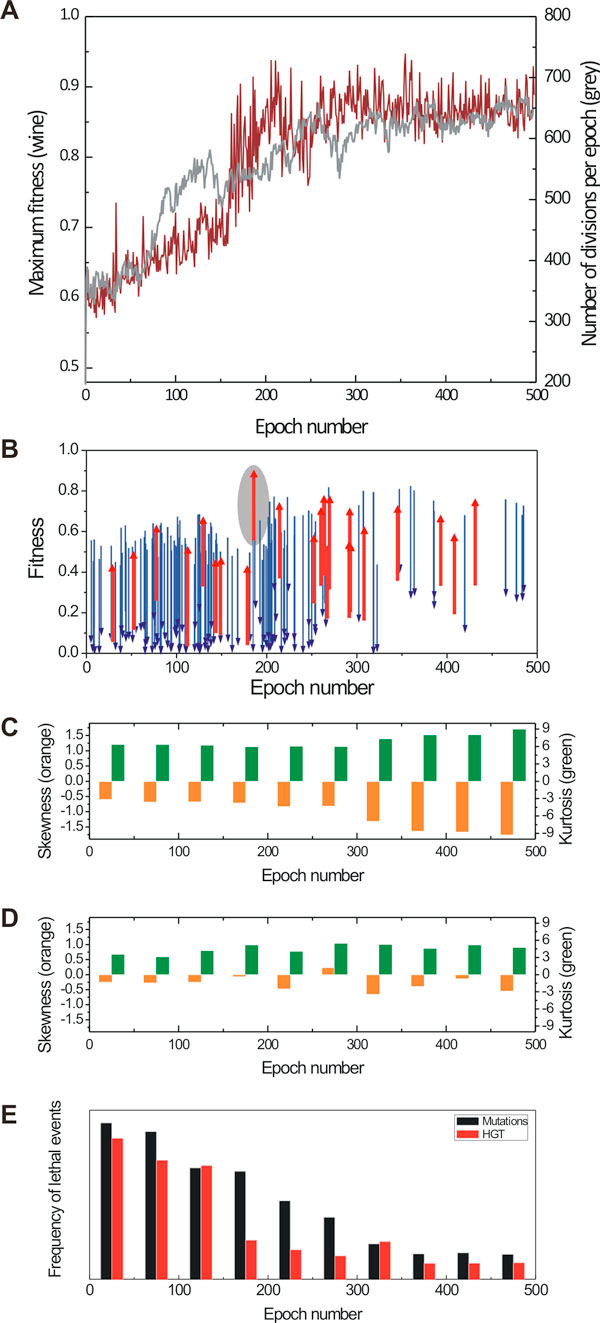
Results: Large-scale simulations reveal a distinct signature of the Distribution of Fitness Effect (DFE) for HGT events: during evolution, while mutation fitness effects become more negative and neutral, HGT events result in a balanced effect distribution. In either case, lethal events are significantly decreased during evolution (33.0% to 3.2%), a clear indication of mutational robustness. Interestingly, evolution was accelerated when populations were exposed to correlated environments of increasing complexity, especially in the presence of HGT, a phenomenon that warrants further investigation. High HGT rates were found to be disruptive, while the average transferred fragment size was linked to functional module size in the underlying biological network. Network analysis reveals that HGT results in larger regulatory networks, but with the same sparsity level as those evolved in its absence. Observed phenotypic variability and co-existing solutions were traced to individual gain/loss of function events, while subsequent re-wiring after fragment integration was necessary for complex traits to emerge.
Figures
Similar articles
-
Guided evolution of in silico microbial populations in complex environments accelerates evolutionary rates through a step-wise adaptation.BMC Bioinformatics. 2012 Jun 25;13 Suppl 10(Suppl 10):S10. doi: 10.1186/1471-2105-13-S10-S10. BMC Bioinformatics. 2012. PMID: 22759415 Free PMC article.
-
Algorithms for computing parsimonious evolutionary scenarios for genome evolution, the last universal common ancestor and dominance of horizontal gene transfer in the evolution of prokaryotes.BMC Evol Biol. 2003 Jan 6;3:2. doi: 10.1186/1471-2148-3-2. Epub 2003 Jan 6. BMC Evol Biol. 2003. PMID: 12515582 Free PMC article.
-
The advantages and disadvantages of horizontal gene transfer and the emergence of the first species.Biol Direct. 2011 Jan 3;6:1. doi: 10.1186/1745-6150-6-1. Biol Direct. 2011. PMID: 21199581 Free PMC article.
-
Exploring the costs of horizontal gene transfer.Trends Ecol Evol. 2013 Aug;28(8):489-95. doi: 10.1016/j.tree.2013.04.002. Epub 2013 May 22. Trends Ecol Evol. 2013. PMID: 23706556 Review.
-
Horizontal gene transfer and adaptive evolution in bacteria.Nat Rev Microbiol. 2022 Apr;20(4):206-218. doi: 10.1038/s41579-021-00650-4. Epub 2021 Nov 12. Nat Rev Microbiol. 2022. PMID: 34773098 Review.
Cited by
-
Extreme Deviations from Expected Evolutionary Rates in Archaeal Protein Families.Genome Biol Evol. 2017 Oct 1;9(10):2791-2811. doi: 10.1093/gbe/evx189. Genome Biol Evol. 2017. PMID: 28985292 Free PMC article.
-
Decoupled genomic elements and the evolution of partner quality in nitrogen-fixing rhizobia.Ecol Evol. 2016 Jan 28;6(5):1317-27. doi: 10.1002/ece3.1953. eCollection 2016 Mar. Ecol Evol. 2016. PMID: 27087920 Free PMC article.
-
Inferring microbial interaction network from microbiome data using RMN algorithm.BMC Syst Biol. 2015 Sep 4;9:54. doi: 10.1186/s12918-015-0199-2. BMC Syst Biol. 2015. PMID: 26337930 Free PMC article.
-
A flood-based information flow analysis and network minimization method for gene regulatory networks.BMC Bioinformatics. 2013 Apr 24;14:137. doi: 10.1186/1471-2105-14-137. BMC Bioinformatics. 2013. PMID: 23617932 Free PMC article.
