

N-linked glycosylation of GP5 of porcine reproductive and respiratory syndrome virus is critically important for virus replication in vivo
- PMID: 22761373
- PMCID: PMC3446624
- DOI: 10.1128/JVI.07067-11
N-linked glycosylation of GP5 of porcine reproductive and respiratory syndrome virus is critically important for virus replication in vivo
Abstract
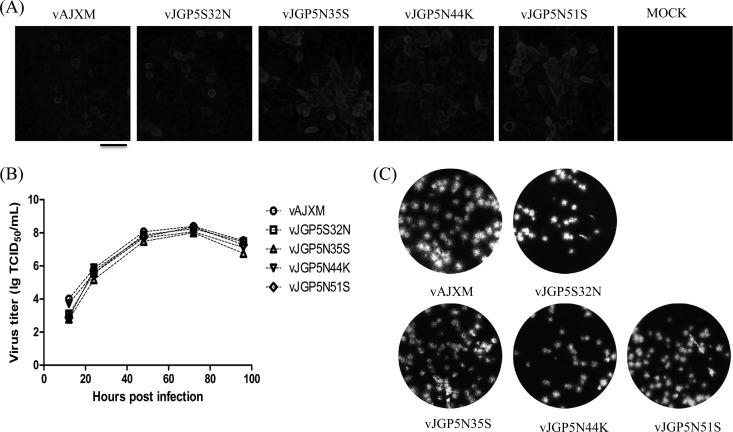
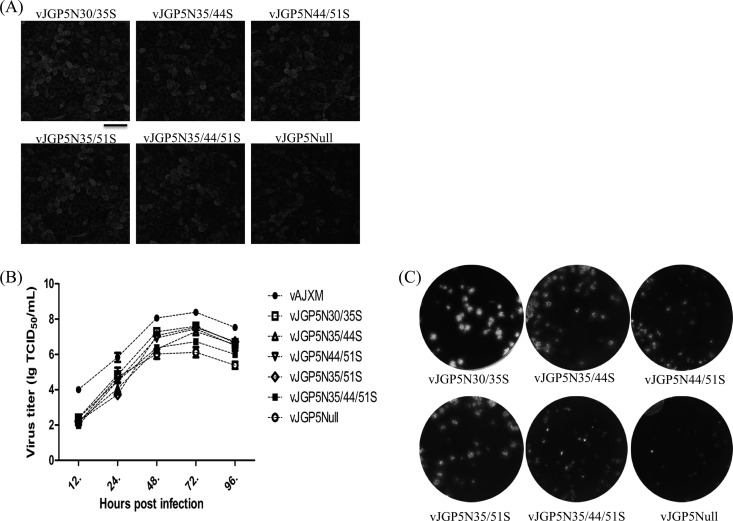
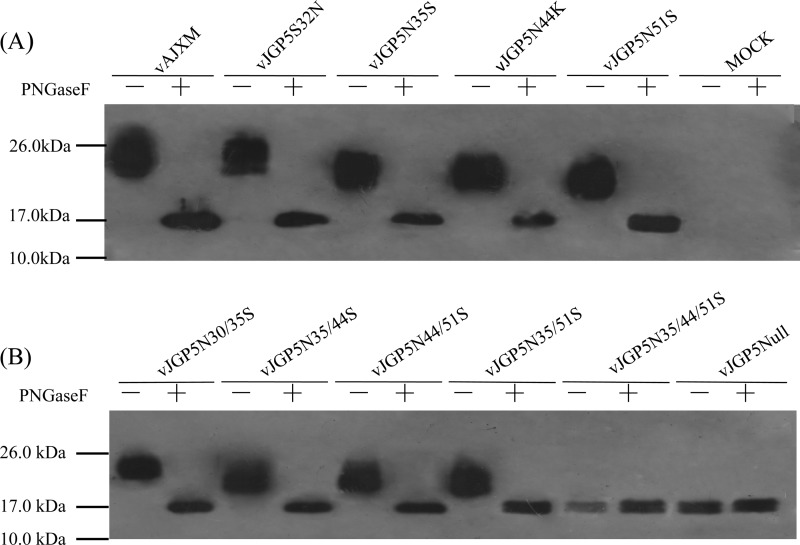
It has been proposed that the N-linked glycan addition at certain sites in GP5 of porcine reproductive and respiratory syndrome virus (PRRSV) is important for production of infectious viruses and viral infectivity. However, such specific N-linked glycosylation sites do not exist in some field PRRSV isolates. This implies that the existence of GP5-associated glycan per se is not vital to the virus life cycle. In this study, we found that mutation of individual glycosylation sites at N30, N35, N44, and N51 in GP5 did not affect virus infectivity in cultured cells. However, the mutants carrying multiple mutations at N-linked glycosylation sites in GP5 had significantly reduced virus yields compared with the wild-type (wt) virus. As a result, no viremia and antibody response were detected in piglets that were injected with a mutant without all N-linked glycans in GP5. These results suggest that the N-linked glycosylation of GP5 is critically important for virus replication in vivo. The study also showed that removal of N44-linked glycan from GP5 increased the sensitivity of mutant virus to convalescent-phase serum samples but did not elicit a high-level neutralizing antibody response to wt PRRSV. The results obtained from the present study have made significant contributions to better understanding the importance of glycosylation of GP5 in the biology of PRRSV.
Figures

Similar articles
-
Influence of N-linked glycosylation of porcine reproductive and respiratory syndrome virus GP5 on virus infectivity, antigenicity, and ability to induce neutralizing antibodies.J Virol. 2006 Apr;80(8):3994-4004. doi: 10.1128/JVI.80.8.3994-4004.2006. J Virol. 2006. PMID: 16571816 Free PMC article.
-
Neutralizing antibody responses of pigs infected with natural GP5 N-glycan mutants of porcine reproductive and respiratory syndrome virus.Viral Immunol. 2006 Summer;19(2):294-304. doi: 10.1089/vim.2006.19.294. Viral Immunol. 2006. PMID: 16817772
-
Influence of porcine reproductive and respiratory syndrome virus GP5 glycoprotein N-linked glycans on immune responses in mice.Virus Genes. 2007 Dec;35(3):663-71. doi: 10.1007/s11262-007-0131-y. Epub 2007 Aug 2. Virus Genes. 2007. PMID: 17671839
-
GP5 of porcine reproductive and respiratory syndrome virus (PRRSV) as a target for homologous and broadly neutralizing antibodies.Vet Microbiol. 2017 Sep;209:90-96. doi: 10.1016/j.vetmic.2017.04.016. Epub 2017 Apr 29. Vet Microbiol. 2017. PMID: 28528961 Review.
-
Research Progress on Glycoprotein 5 of Porcine Reproductive and Respiratory Syndrome Virus.Animals (Basel). 2023 Feb 23;13(5):813. doi: 10.3390/ani13050813. Animals (Basel). 2023. PMID: 36899670 Free PMC article. Review.
Cited by
-
Impact of genetic variation and geographic distribution of porcine reproductive and respiratory syndrome virus on infectivity and pig growth.BMC Vet Res. 2013 Mar 27;9:58. doi: 10.1186/1746-6148-9-58. BMC Vet Res. 2013. PMID: 23537091 Free PMC article.
-
Glyceraldehyde-3-Phosphate Dehydrogenase Restricted in Cytoplasmic Location by Viral GP5 Facilitates Porcine Reproductive and Respiratory Syndrome Virus Replication via Its Glycolytic Activity.J Virol. 2021 Aug 25;95(18):e0021021. doi: 10.1128/JVI.00210-21. Epub 2021 Aug 25. J Virol. 2021. PMID: 34160254 Free PMC article.
-
Optimized protocol for expression and purification of membrane-bound PglB, a bacterial oligosaccharyl transferase.Protein Expr Purif. 2013 Jun;89(2):241-50. doi: 10.1016/j.pep.2013.04.001. Epub 2013 Apr 12. Protein Expr Purif. 2013. PMID: 23583934 Free PMC article.
-
Evolutionary Dynamics of Type 2 Porcine Reproductive and Respiratory Syndrome Virus by Whole-Genome Analysis.Viruses. 2021 Dec 9;13(12):2469. doi: 10.3390/v13122469. Viruses. 2021. PMID: 34960738 Free PMC article.
-
Role of N-linked glycosylation in porcine reproductive and respiratory syndrome virus (PRRSV) infection.J Gen Virol. 2024 May;105(5):001994. doi: 10.1099/jgv.0.001994. J Gen Virol. 2024. PMID: 38776134 Free PMC article.
References
-
- Allende R, et al. 1999. North American and European porcine reproductive and respiratory syndrome viruses differ in non-structural protein coding regions. J. Gen. Virol. 80:307–315 - PubMed
-
- Chen Z, Li K, Plagemann PG. 2000. Neuropathogenicity and sensitivity to antibody neutralization of lactate dehydrogenase-elevating virus are determined by polylactosaminoglycan chains on the primary envelope glycoprotein. Virology 266:88–98 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
