

Rotavirus VP8*: phylogeny, host range, and interaction with histo-blood group antigens
- PMID: 22761376
- PMCID: PMC3446626
- DOI: 10.1128/JVI.00979-12
Rotavirus VP8*: phylogeny, host range, and interaction with histo-blood group antigens
Abstract
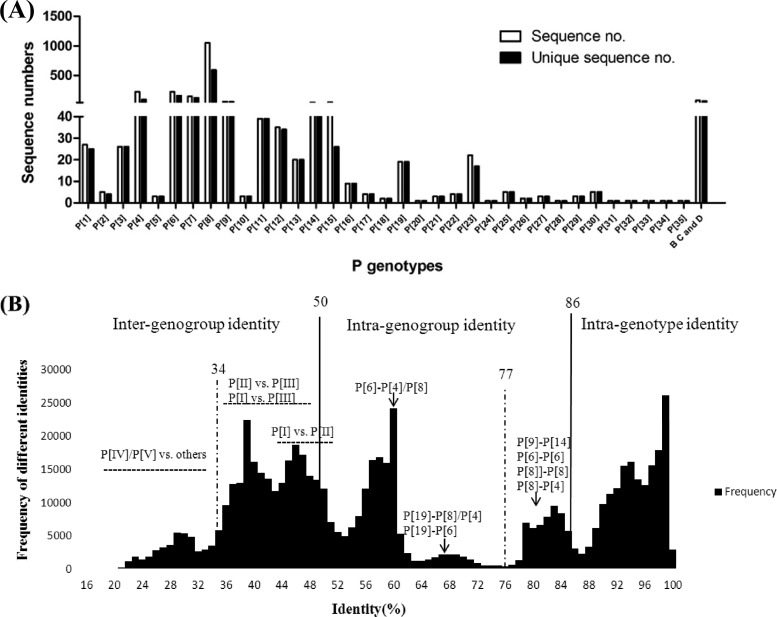
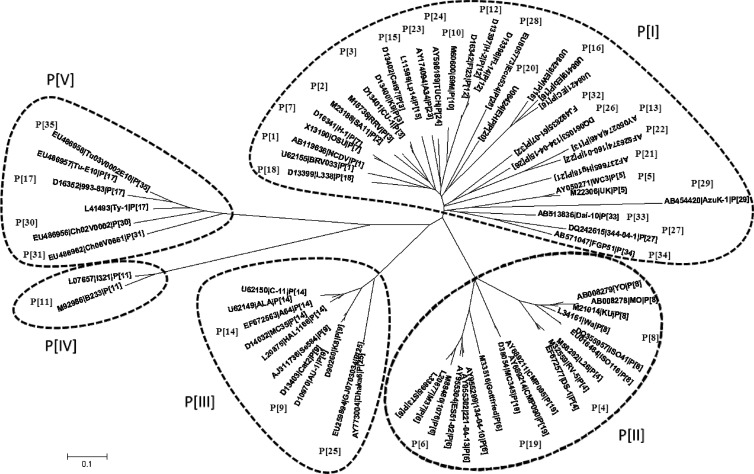
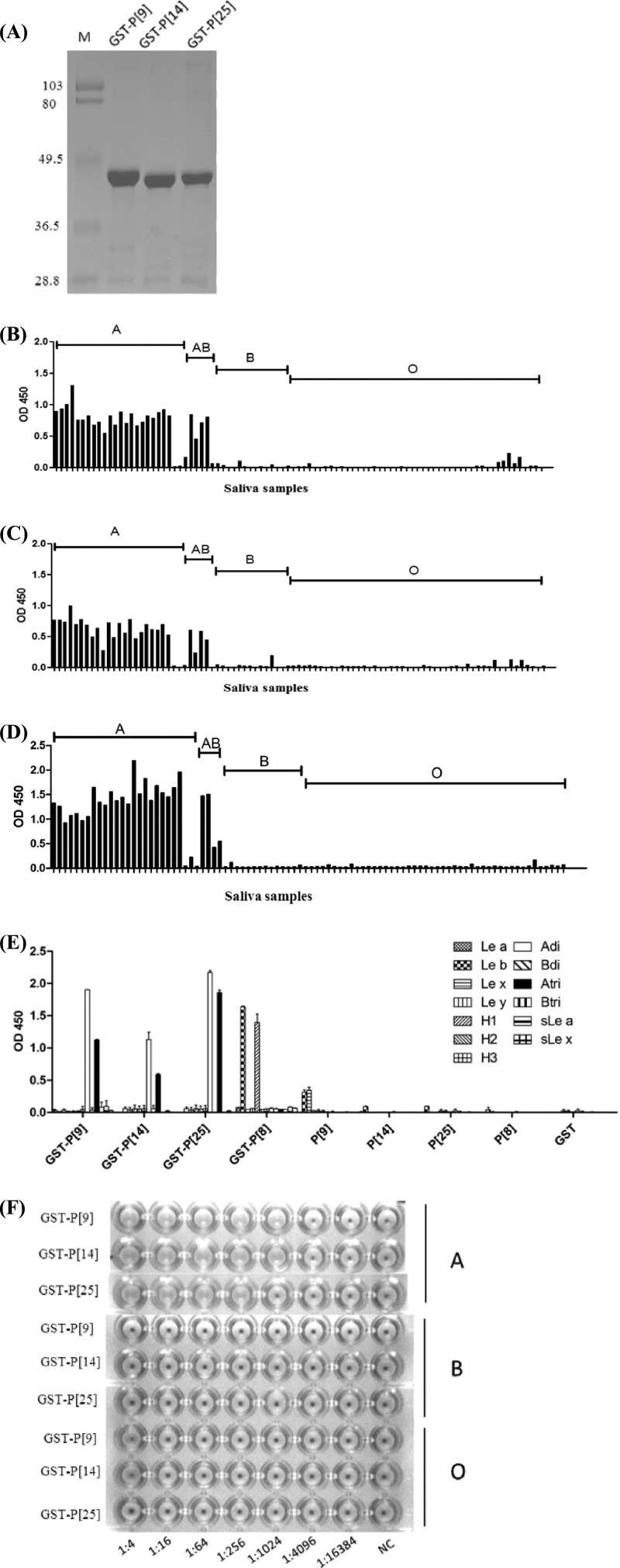
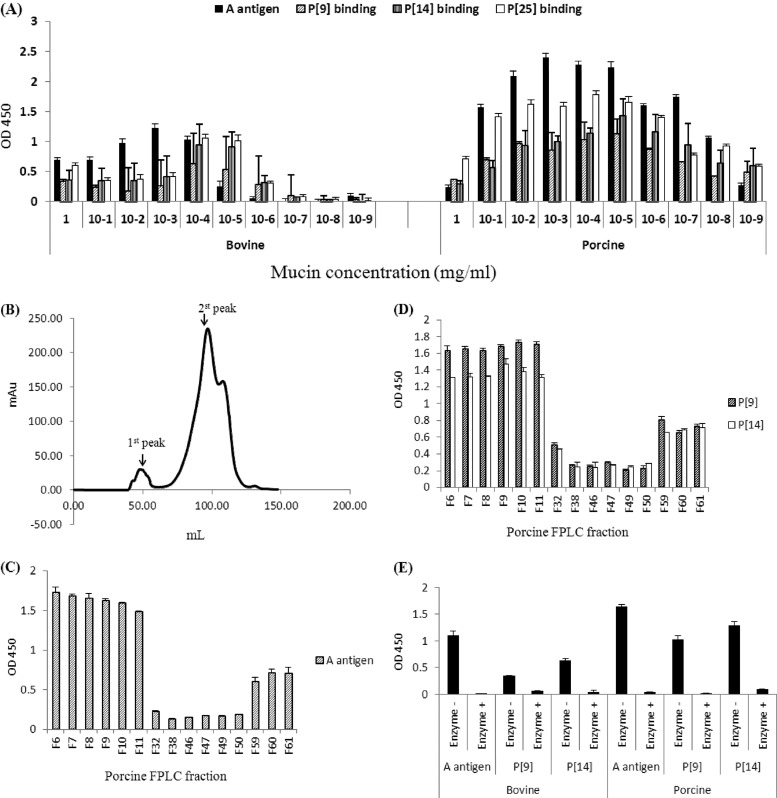
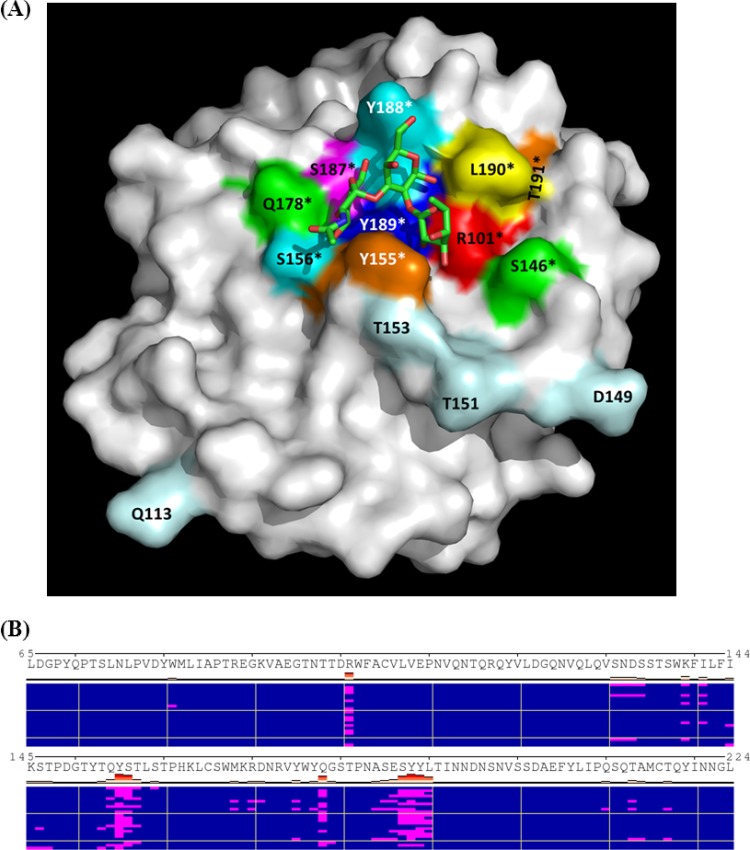
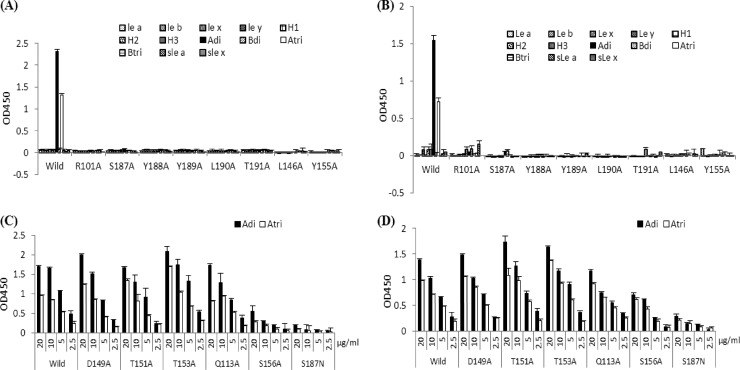
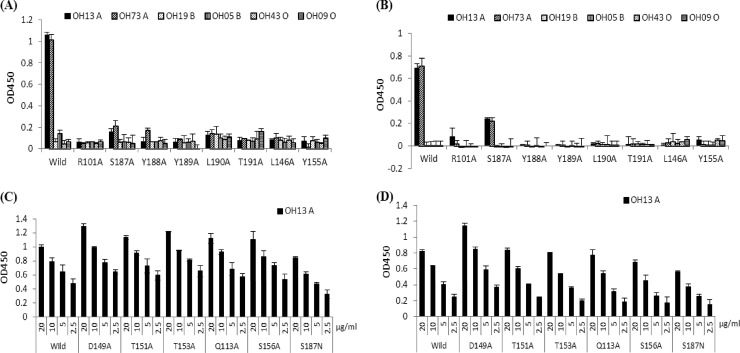
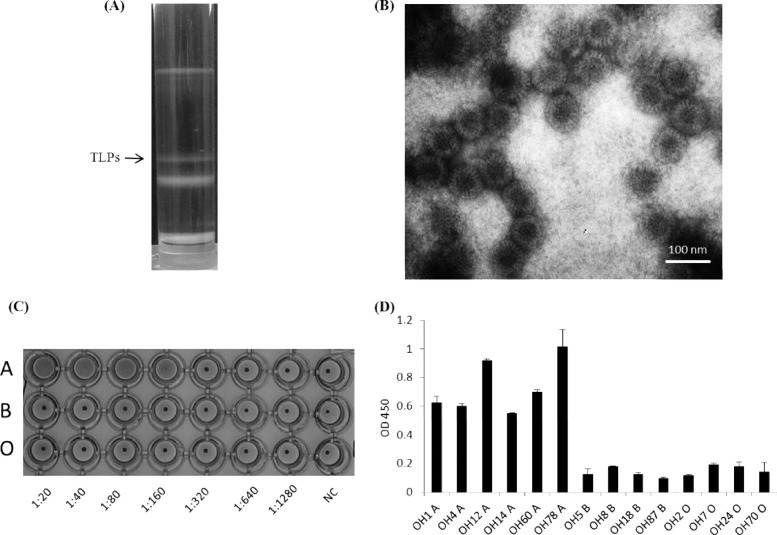
The distal portion of rotavirus (RV) VP4 spike protein (VP8*) is implicated in binding to cellular receptors, thereby facilitating viral attachment and entry. While VP8* of some animal RVs engage sialic acid, human RVs often attach to and enter cells in a sialic acid-independent manner. A recent study demonstrated that the major human RVs (P[4], P[6], and P[8]) recognize human histo-blood group antigens (HBGAs). In this study, we performed a phylogenetic analysis of RVs and showed further variations of RV interaction with HBGAs. On the basis of the VP8* sequences, RVs are grouped into five P genogroups (P[I] to P[V]), of which P[I], P[IV], and P[V] mainly infect animals, P[II] infects humans, and P[III] infects both animals and humans. The sialic acid-dependent RVs (P[1], P[2], P[3], and P[7]) form a subcluster within P[I], while all three major P genotypes of human RVs (P[4], P[6], and P[8]) are clustered in P[II]. We then characterized three human RVs (P[9], P[14], and P[25]) in P[III] and observed a new pattern of binding to the type A antigen which is distinct from that of the P[II] RVs. The binding was demonstrated by hemagglutination and saliva binding assay using recombinant VP8* and native RVs. Homology modeling and mutagenesis study showed that the locations of the carbohydrate binding interfaces are shared with the sialic acid-dependent RVs, although different amino acids are involved. The P[III] VP8* proteins also bind the A antigens of the porcine and bovine mucins, suggesting the A antigen as a possible factor for cross-species transmission of RVs. Our study suggests that HBGAs play an important role in RV infection and evolution.
Figures
References
-
- Beards G, Graham C. 1995. Temporal distribution of rotavirus G-serotypes in the West Midlands region of the United Kingdom, 1983–1994. J. Diarrhoeal Dis. Res. 13:235–237 - PubMed
-
- Bishop JR, Gagneux P. 2007. Evolution of carbohydrate antigens—microbial forces shaping host glycomes? Glycobiology 17:23R–34R - PubMed
-
- Castello AA, et al. 2009. Characterization of genotype P[9]G12 rotavirus strains from Argentina: high similarity with Japanese and Korean G12 strains J. Med. Virol. 81:371–381 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
