

Adaptive changes in alphavirus mRNA translation allowed colonization of vertebrate hosts
- PMID: 22761388
- PMCID: PMC3416173
- DOI: 10.1128/JVI.01114-12
Adaptive changes in alphavirus mRNA translation allowed colonization of vertebrate hosts
Abstract
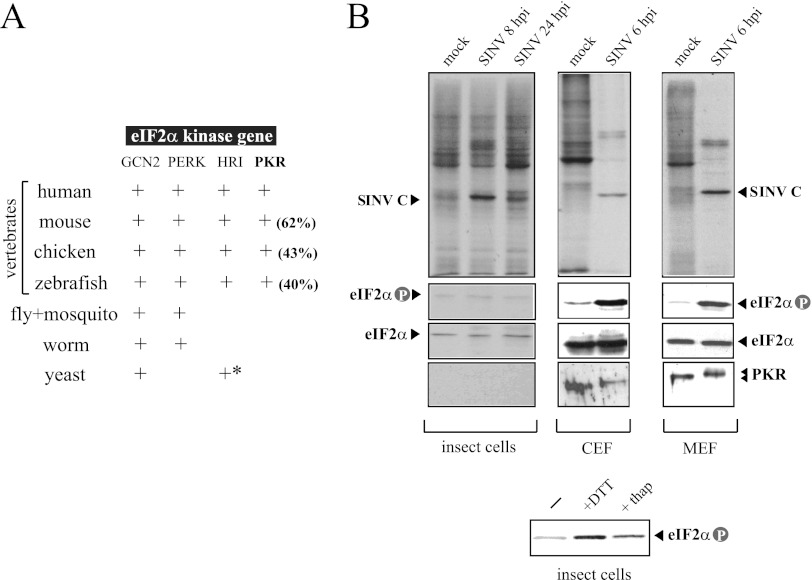
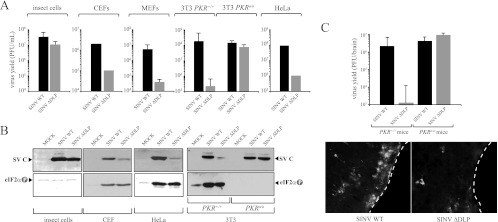
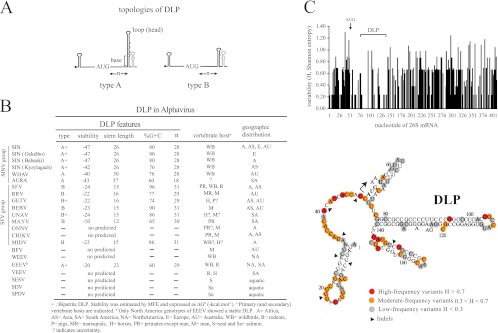
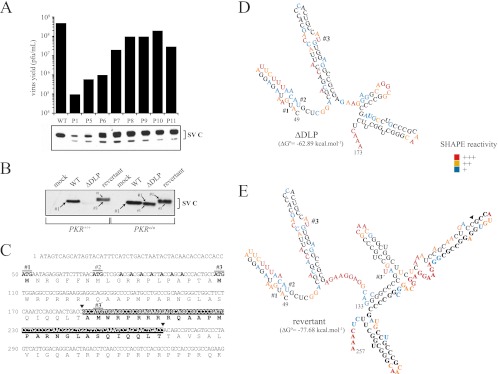
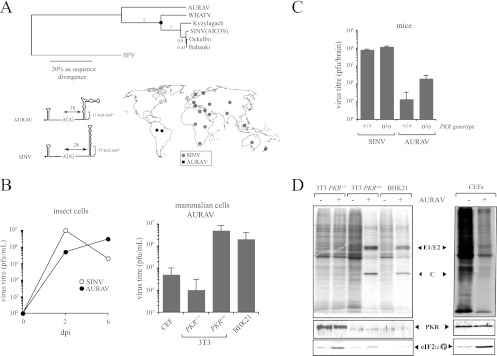
Members of the Alphavirus genus are arboviruses that alternate replication in mosquitoes and vertebrate hosts. In vertebrate cells, the alphavirus resists the activation of antiviral RNA-activated protein kinase (PKR) by the presence of a prominent RNA structure (downstream loop [DLP]) located in viral 26S transcripts, which allows an eIF2-independent translation initiation of these mRNAs. This article shows that DLP structure is essential for replication of Sindbis virus (SINV) in vertebrate cell lines and animals but is dispensable for replication in insect cells, where no ortholog of the vertebrate PKR gene has been found. Sequence comparisons and structural RNA analysis revealed the evolutionary conservation of DLP in SINV and predicted the existence of equivalent DLP structures in many members of the Alphavirus genus. A mutant SINV lacking the DLP structure evolved in murine cells to recover a wild-type phenotype by creating an alternative structure in the RNA that restored the translational independence for eIF2. Genetic, phylogenetic, and biochemical data presented here support an evolutionary scenario for the natural history of alphaviruses, in which the acquisition of DLP structure in their mRNAs probably allowed the colonization of vertebrate host and the consequent geographic expansion of some of these viruses worldwide.
Figures
Similar articles
-
Translational Control of Alphavirus-Host Interactions: Implications in Viral Evolution, Tropism and Antiviral Response.Viruses. 2024 Jan 30;16(2):205. doi: 10.3390/v16020205. Viruses. 2024. PMID: 38399981 Free PMC article. Review.
-
An RNA trapping mechanism in Alphavirus mRNA promotes ribosome stalling and translation initiation.Nucleic Acids Res. 2016 May 19;44(9):4368-80. doi: 10.1093/nar/gkw172. Epub 2016 Mar 16. Nucleic Acids Res. 2016. PMID: 26984530 Free PMC article.
-
Naturally Occurring and Engineered Alphaviruses Sensitive to Double-Stranded-RNA-Activated Protein Kinase Show Restricted Translation in Mammalian Cells, Increased Sensitivity to Interferon, and Marked Oncotropism.J Virol. 2020 Jan 17;94(3):e01630-19. doi: 10.1128/JVI.01630-19. Print 2020 Jan 17. J Virol. 2020. PMID: 31723025 Free PMC article.
-
Diversity in viral anti-PKR mechanisms: a remarkable case of evolutionary convergence.PLoS One. 2011 Feb 2;6(2):e16711. doi: 10.1371/journal.pone.0016711. PLoS One. 2011. PMID: 21311764 Free PMC article.
-
The Regulation of Translation in Alphavirus-Infected Cells.Viruses. 2018 Feb 8;10(2):70. doi: 10.3390/v10020070. Viruses. 2018. PMID: 29419763 Free PMC article. Review.
Cited by
-
Translational Control of Alphavirus-Host Interactions: Implications in Viral Evolution, Tropism and Antiviral Response.Viruses. 2024 Jan 30;16(2):205. doi: 10.3390/v16020205. Viruses. 2024. PMID: 38399981 Free PMC article. Review.
-
Improvement of In Vivo Expression of Genes Delivered by Self-Amplifying RNA Using Vaccinia Virus Immune Evasion Proteins.Hum Gene Ther. 2017 Dec;28(12):1138-1146. doi: 10.1089/hum.2017.121. Epub 2017 Sep 6. Hum Gene Ther. 2017. PMID: 28877647 Free PMC article.
-
NF-κB Activation Promotes Alphavirus Replication in Mature Neurons.J Virol. 2019 Nov 26;93(24):e01071-19. doi: 10.1128/JVI.01071-19. Print 2019 Dec 15. J Virol. 2019. PMID: 31554691 Free PMC article.
-
Salmonid alphavirus replication in mosquito cells: towards a novel vaccine production system.Microb Biotechnol. 2014 Sep;7(5):480-4. doi: 10.1111/1751-7915.12100. Epub 2014 Jan 14. Microb Biotechnol. 2014. PMID: 24418177 Free PMC article.
-
An RNA trapping mechanism in Alphavirus mRNA promotes ribosome stalling and translation initiation.Nucleic Acids Res. 2016 May 19;44(9):4368-80. doi: 10.1093/nar/gkw172. Epub 2016 Mar 16. Nucleic Acids Res. 2016. PMID: 26984530 Free PMC article.
References
-
- Aarti I, Rajesh K, Ramaiah KV. 2010. Phosphorylation of eIF2 alpha in Sf9 cells: a stress, survival and suicidal signal. Apoptosis 15:679–692 - PubMed
-
- Abraham N, et al. 1999. Characterization of transgenic mice with targeted disruption of the catalytic domain of the double-stranded RNA-dependent protein kinase, PKR. J. Biol. Chem. 274:5953–5962 - PubMed
-
- Binder GK, Griffin DE. 2001. Interferon-gamma-mediated site-specific clearance of alphavirus from CNS neurons. Science 293:303–306 - PubMed
-
- Choi HK, et al. 1991. Structure of Sindbis virus core protein reveals a chymotrypsin-like serine proteinase and the organization of the virion. Nature 354:37–43 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
