

Bone marrow oxytocin mediates the anabolic action of estrogen on the skeleton
- PMID: 22761429
- PMCID: PMC3436530
- DOI: 10.1074/jbc.M112.365049
Bone marrow oxytocin mediates the anabolic action of estrogen on the skeleton
Abstract
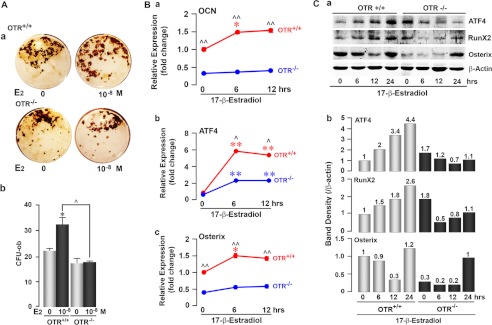
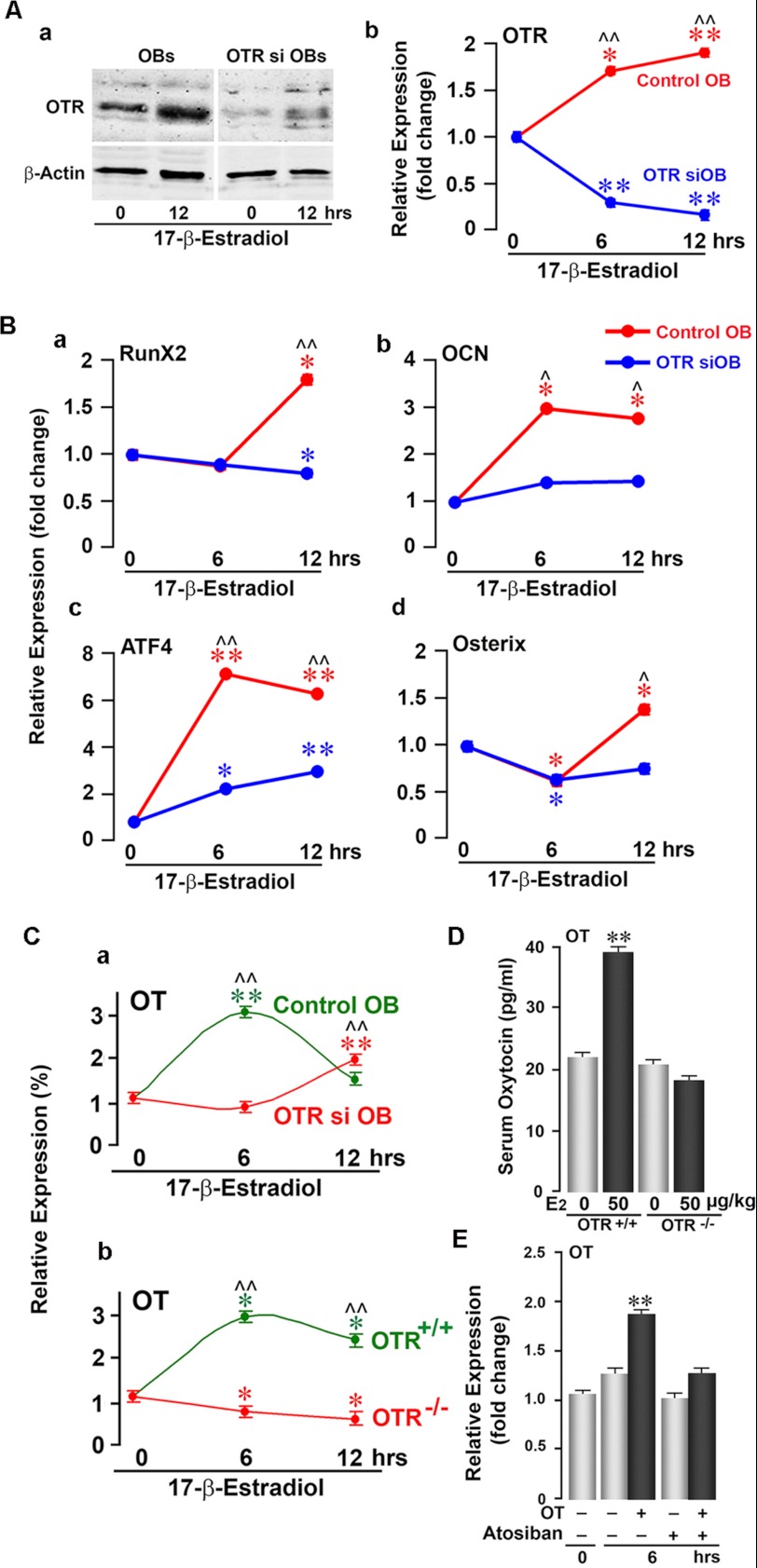
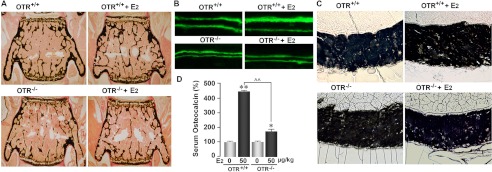
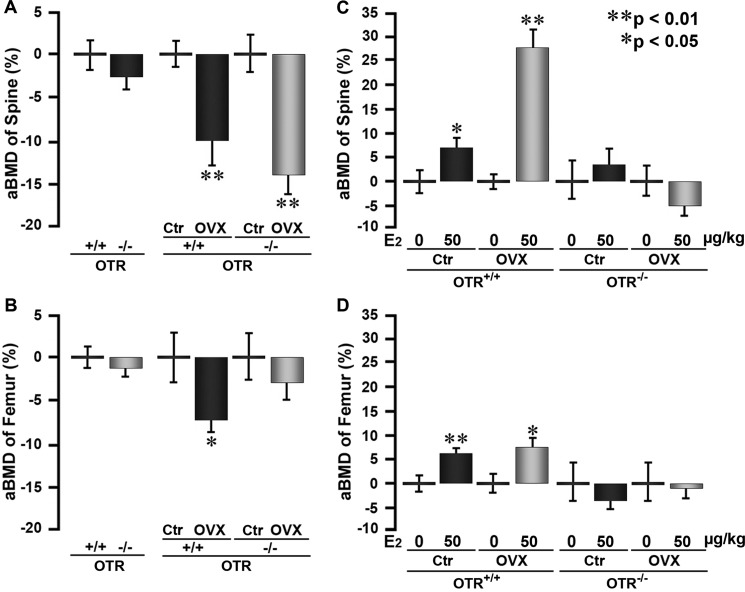
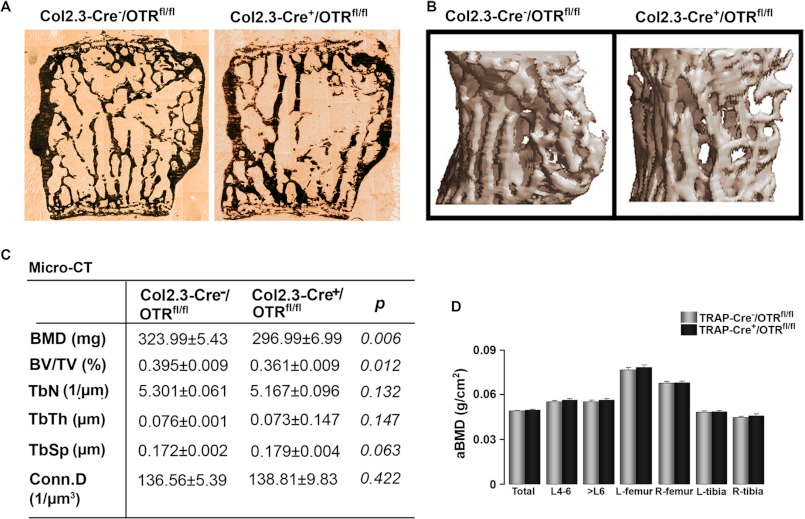
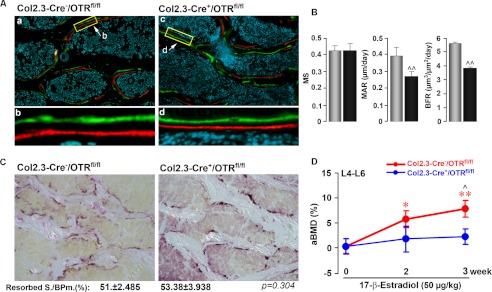
Estrogen uses two mechanisms to exert its effect on the skeleton: it inhibits bone resorption by osteoclasts and, at higher doses, can stimulate bone formation. Although the antiresorptive action of estrogen arises from the inhibition of the MAPK JNK, the mechanism of its effect on the osteoblast remains unclear. Here, we report that the anabolic action of estrogen in mice occurs, at least in part, through oxytocin (OT) produced by osteoblasts in bone marrow. We show that the absence of OT receptors (OTRs) in OTR(-/-) osteoblasts or attenuation of OTR expression in silenced cells inhibits estrogen-induced osteoblast differentiation, transcription factor up-regulation, and/or OT production in vitro. In vivo, OTR(-/-) mice, known to have a bone formation defect, fail to display increases in trabecular bone volume, cortical thickness, and bone formation in response to estrogen. Furthermore, osteoblast-specific Col2.3-Cre(+)/OTR(fl/fl) mice, but not TRAP-Cre(+)/OTR(fl/fl) mice, mimic the OTR(-/-) phenotype and also fail to respond to estrogen. These data attribute the phenotype of OTR deficiency to an osteoblastic rather than an osteoclastic defect. Physiologically, feed-forward OT release in bone marrow by a rising estrogen concentration may facilitate rapid skeletal recovery during the latter phases of lactation.
Figures
References
-
- Breton C., Di Scala-Guenot D., Zingg H. H. (2001) Oxytocin receptor gene expression in rat mammary gland: structural characterization and regulation. J. Mol. Endocrinol. 27, 175–189 - PubMed
-
- Blanks A. M., Thornton S. (2003) The role of oxytocin in parturition. BJOG 110, 46–51 - PubMed
-
- Kovacs C. S. (2001) Calcium and bone metabolism in pregnancy and lactation. J. Clin. Endocrinol. Metab. 86, 2344–2348 - PubMed
-
- VanHouten J. N., Wysolmerski J. J. (2003) Low estrogen and high parathyroid hormone-related peptide levels contribute to accelerated bone resorption and bone loss in lactating mice. Endocrinology 144, 5521–5529 - PubMed
-
- Michalakis K., Peitsidis P., Ilias I. (2011) Pregnancy- and lactation-associated osteoporosis. Endocr. Regul. 45, 43–47 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
