

G protein activation without a GEF in the plant kingdom
- PMID: 22761582
- PMCID: PMC3386157
- DOI: 10.1371/journal.pgen.1002756
G protein activation without a GEF in the plant kingdom
Abstract
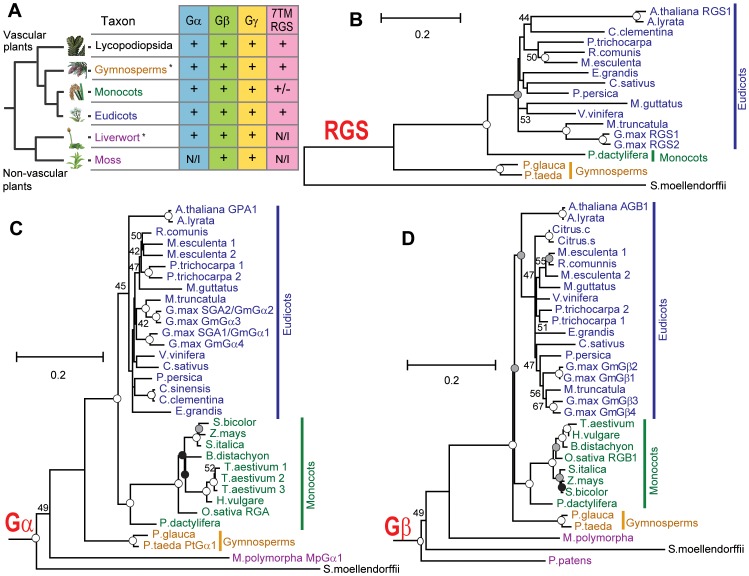
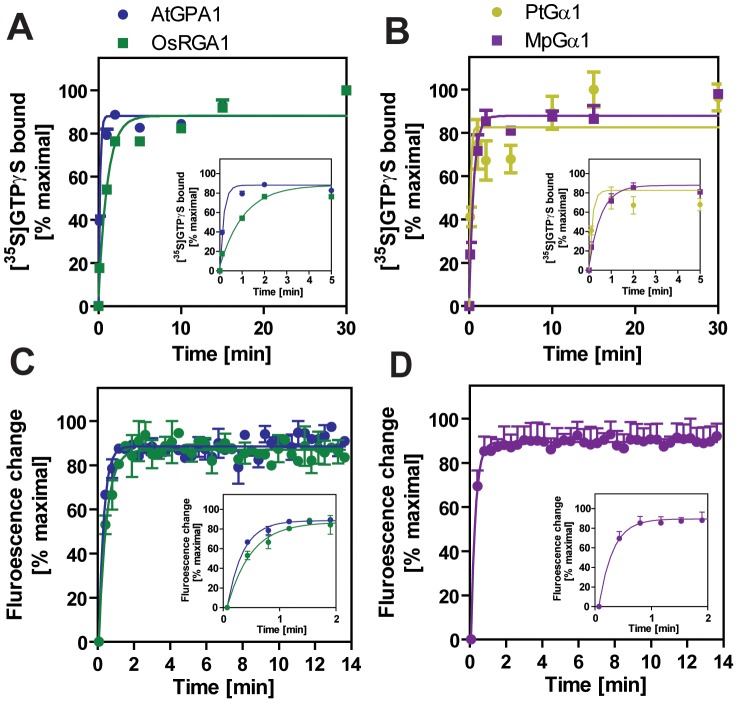
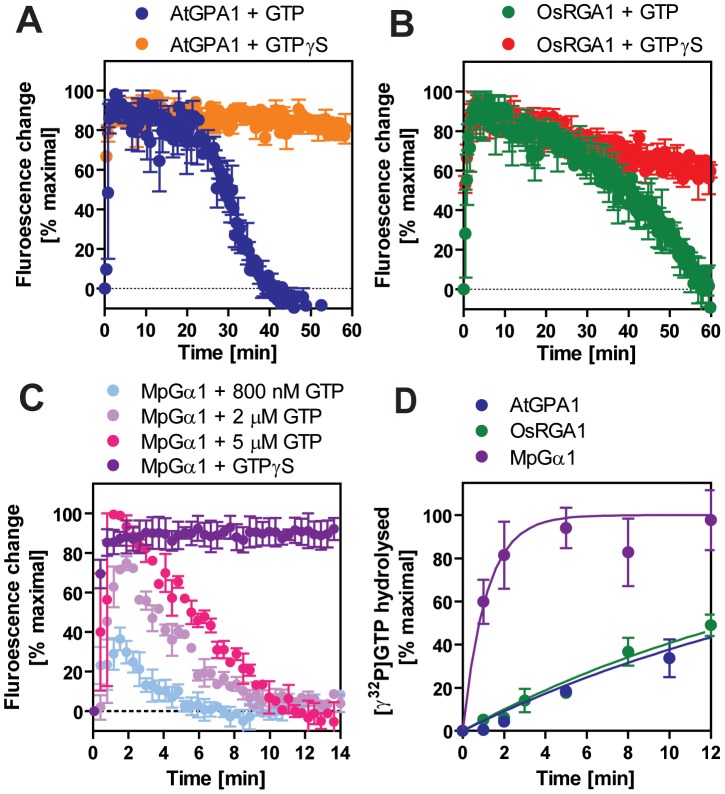
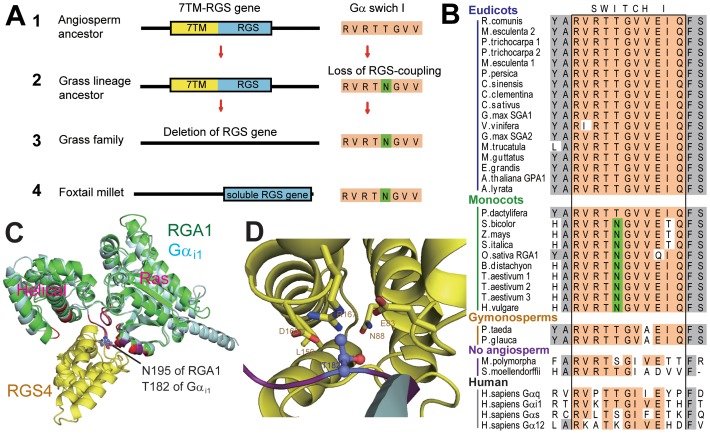
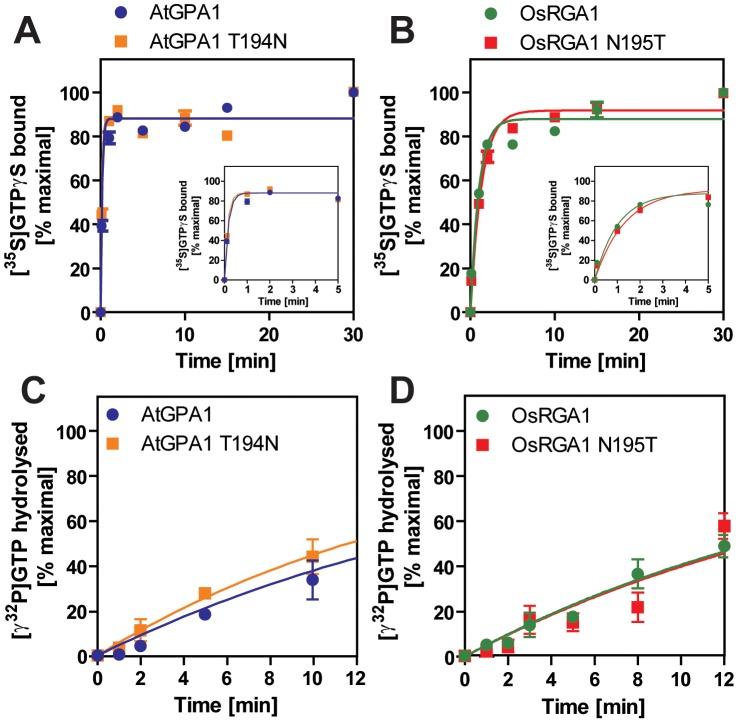
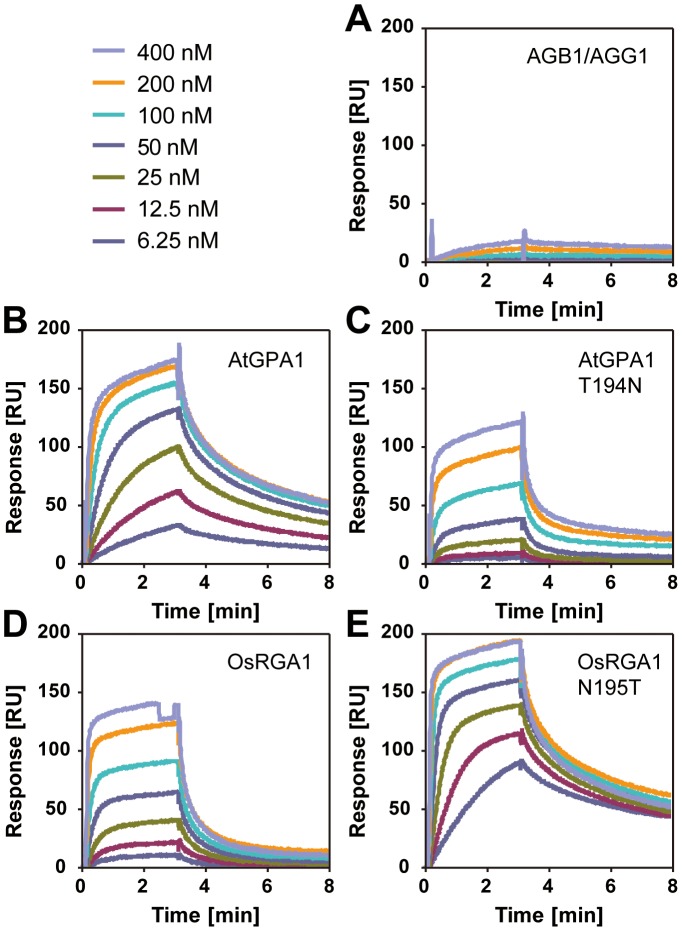
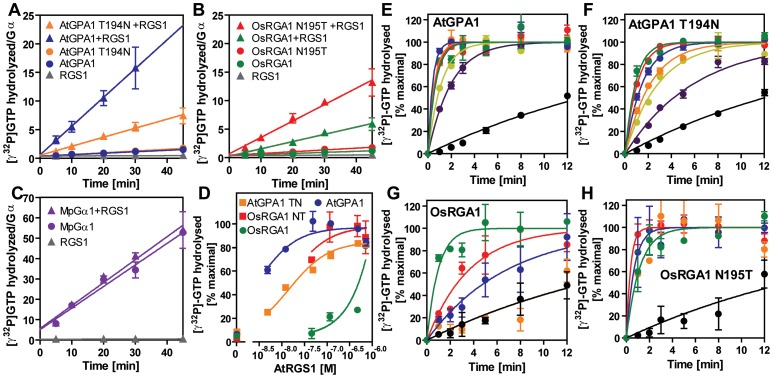
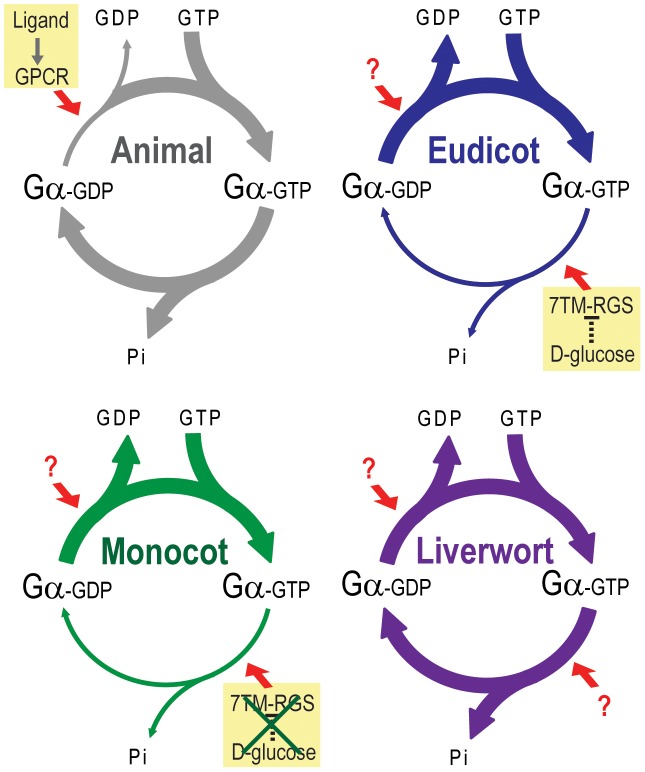
Animal heterotrimeric G proteins are activated by guanine nucleotide exchange factors (GEF), typically seven transmembrane receptors that trigger GDP release and subsequent GTP binding. In contrast, the Arabidopsis thaliana G protein (AtGPA1) rapidly activates itself without a GEF and is instead regulated by a seven transmembrane Regulator of G protein Signaling (7TM-RGS) protein that promotes GTP hydrolysis to reset the inactive (GDP-bound) state. It is not known if this unusual activation is a major and constraining part of the evolutionary history of G signaling in eukaryotes. In particular, it is not known if this is an ancestral form or if this mechanism is maintained, and therefore constrained, within the plant kingdom. To determine if this mode of signal regulation is conserved throughout the plant kingdom, we analyzed available plant genomes for G protein signaling components, and we purified individually the plant components encoded in an informative set of plant genomes in order to determine their activation properties in vitro. While the subunits of the heterotrimeric G protein complex are encoded in vascular plant genomes, the 7TM-RGS genes were lost in all investigated grasses. Despite the absence of a Gα-inactivating protein in grasses, all vascular plant Gα proteins examined rapidly released GDP without a receptor and slowly hydrolyzed GTP, indicating that these Gα are self-activating. We showed further that a single amino acid substitution found naturally in grass Gα proteins reduced the Gα-RGS interaction, and this amino acid substitution occurred before the loss of the RGS gene in the grass lineage. Like grasses, non-vascular plants also appear to lack RGS proteins. However, unlike grasses, one representative non-vascular plant Gα showed rapid GTP hydrolysis, likely compensating for the loss of the RGS gene. Our findings, the loss of a regulatory gene and the retention of the "self-activating" trait, indicate the existence of divergent Gα regulatory mechanisms in the plant kingdom. In the grasses, purifying selection on the regulatory gene was lost after the physical decoupling of the RGS protein and its cognate Gα partner. More broadly these findings show extreme divergence in Gα activation and regulation that played a critical role in the evolution of G protein signaling pathways.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
