

Synaptic vesicle endocytosis
- PMID: 22763746
- PMCID: PMC3428771
- DOI: 10.1101/cshperspect.a005645
Synaptic vesicle endocytosis
Abstract
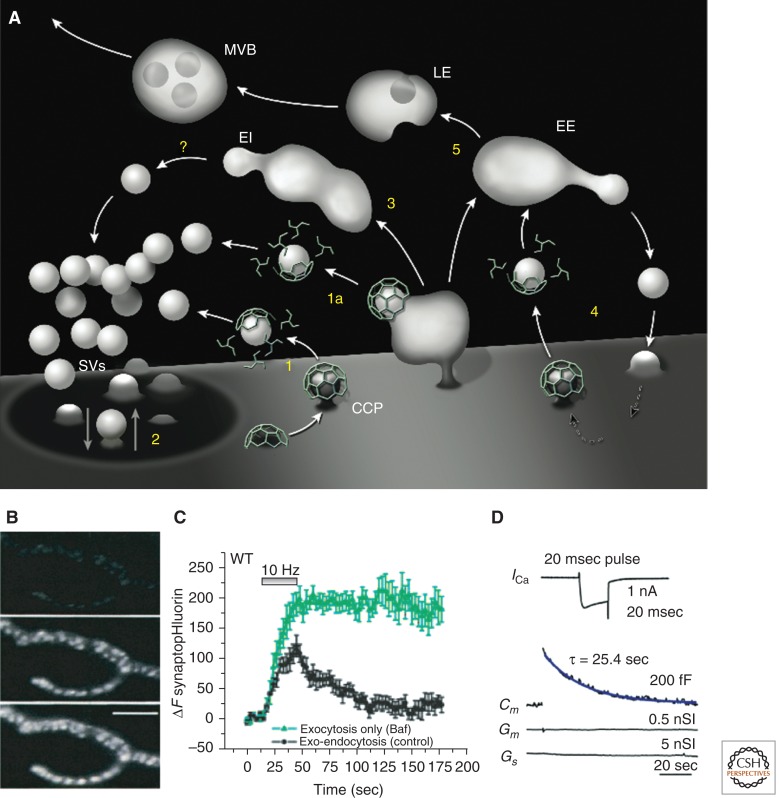
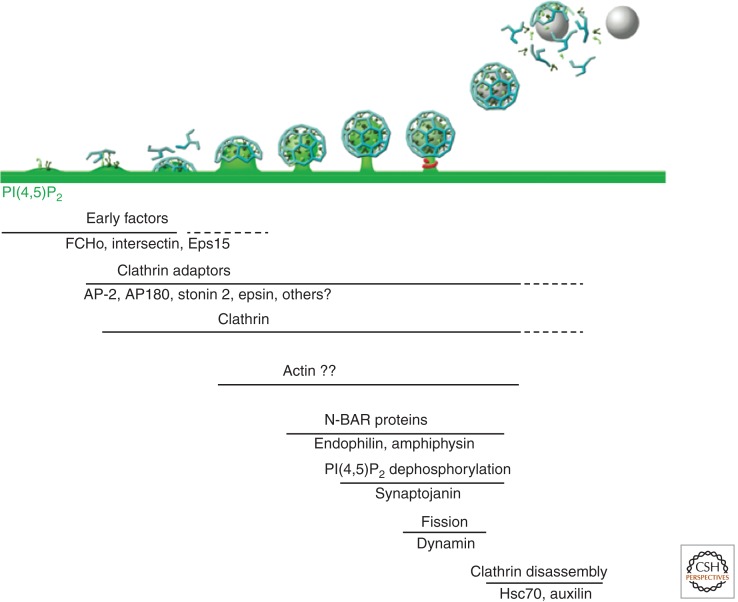
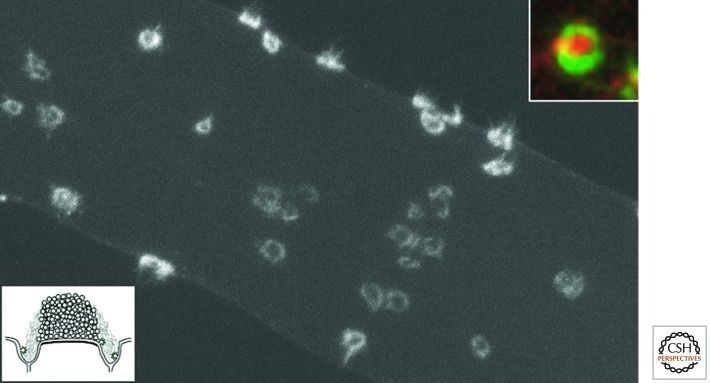
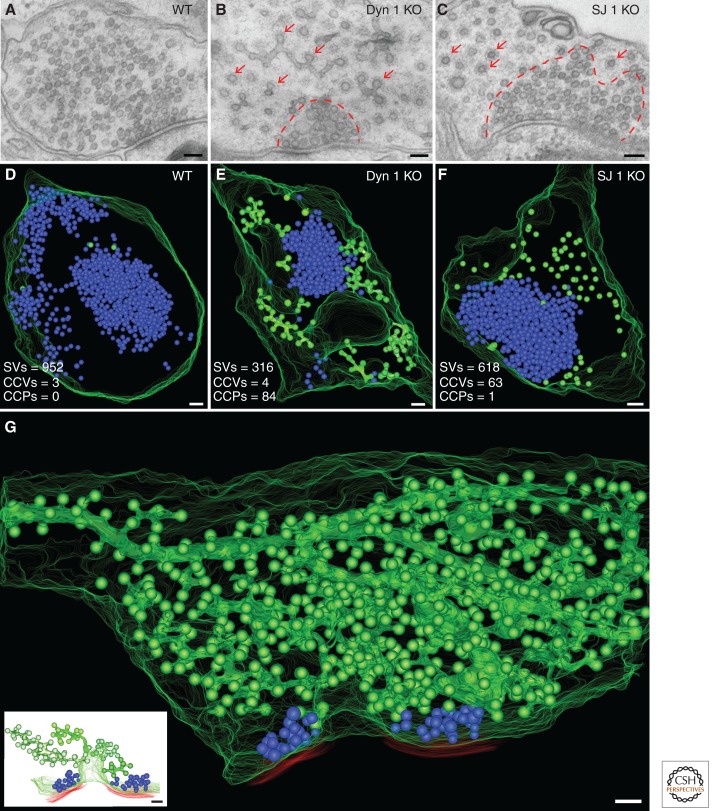
Neurons can sustain high rates of synaptic transmission without exhausting their supply of synaptic vesicles. This property relies on a highly efficient local endocytic recycling of synaptic vesicle membranes, which can be reused for hundreds, possibly thousands, of exo-endocytic cycles. Morphological, physiological, molecular, and genetic studies over the last four decades have provided insight into the membrane traffic reactions that govern this recycling and its regulation. These studies have shown that synaptic vesicle endocytosis capitalizes on fundamental and general endocytic mechanisms but also involves neuron-specific adaptations of such mechanisms. Thus, investigations of these processes have advanced not only the field of synaptic transmission but also, more generally, the field of endocytosis. This article summarizes current information on synaptic vesicle endocytosis with an emphasis on the underlying molecular mechanisms and with a special focus on clathrin-mediated endocytosis, the predominant pathway of synaptic vesicle protein internalization.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources