

Epigenetic regulation of gene expression in keratinocytes
- PMID: 22763788
- PMCID: PMC3650472
- DOI: 10.1038/jid.2012.182
Epigenetic regulation of gene expression in keratinocytes
Abstract
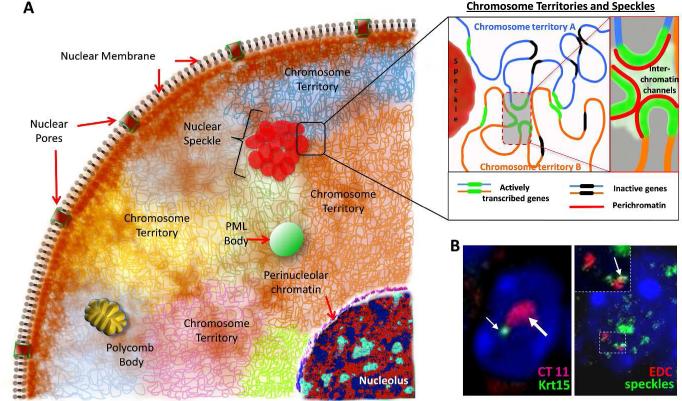
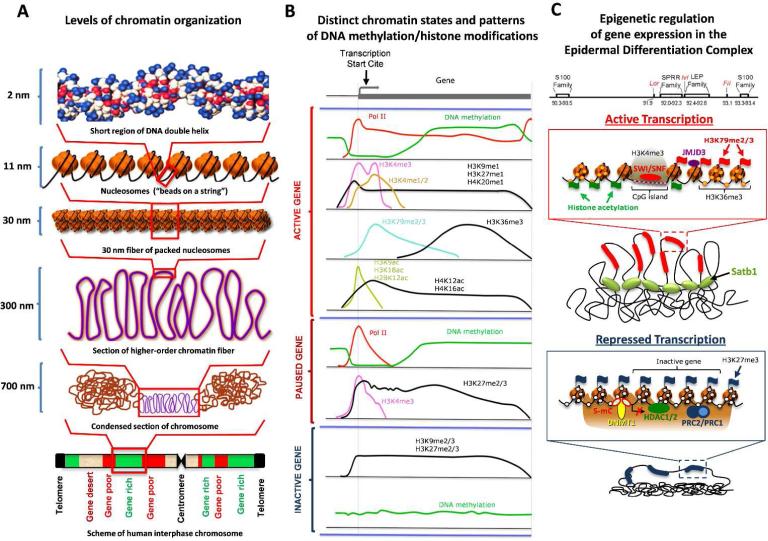
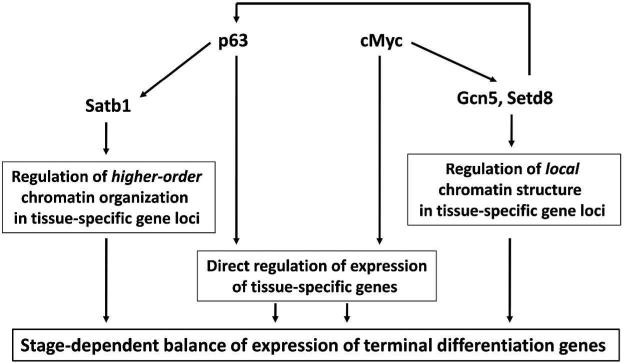
The nucleus is a complex and highly compartmentalized organelle, which undergoes major organization changes during cell differentiation, allowing cells to become specialized and fulfill their functions. During terminal differentiation of the epidermal keratinocytes, the nucleus undergoes a programmed transformation from active status, associated with execution of the genetic programs of cornification and epidermal barrier formation, to a fully inactive condition and becomes a part of the keratinized cells of the cornified layer. Tremendous progress achieved within the past two decades in understanding the biology of the nucleus and epigenetic mechanisms controlling gene expression allowed defining several levels in the regulation of cell differentiation-associated gene expression programs, including an accessibility of the gene regulatory regions to DNA-protein interactions, covalent DNA and histone modifications, and ATP-dependent chromatin remodeling, as well as higher-order chromatin remodeling and nuclear compartmentalization of the genes and transcription machinery. Here, we integrate our current knowledge of the mechanisms controlling gene expression during terminal keratinocyte differentiation with distinct levels of chromatin organization and remodeling. We also propose directions to further explore the role of epigenetic mechanisms and their interactions with other regulatory systems in the control of keratinocyte differentiation in normal and diseased skin.
Figures
References
-
- Allis CD, Berger SL, Cote J, Dent S, Jenuwien T, Kouzarides T, et al. New nomenclature for chromatin-modifying enzymes. Cell. 2007;131:633–6. - PubMed
-
- Amano T, Sagai T, Tanabe H, Mizushina Y, Nakazawa H, Shiroishi T. Chromosomal dynamics at the Shh locus: limb bud-specific differential regulation of competence and active transcription. Dev Cell. 2009;16:47–57. - PubMed
-
- Andrews AJ, Luger K. Nucleosome structure(s) and stability: variations on a theme. Annu Rev Biophys. 2011;40:99–117. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
