

Multiscale mechanisms of cell migration during development: theory and experiment
- PMID: 22764050
- PMCID: PMC3403103
- DOI: 10.1242/dev.081471
Multiscale mechanisms of cell migration during development: theory and experiment
Abstract
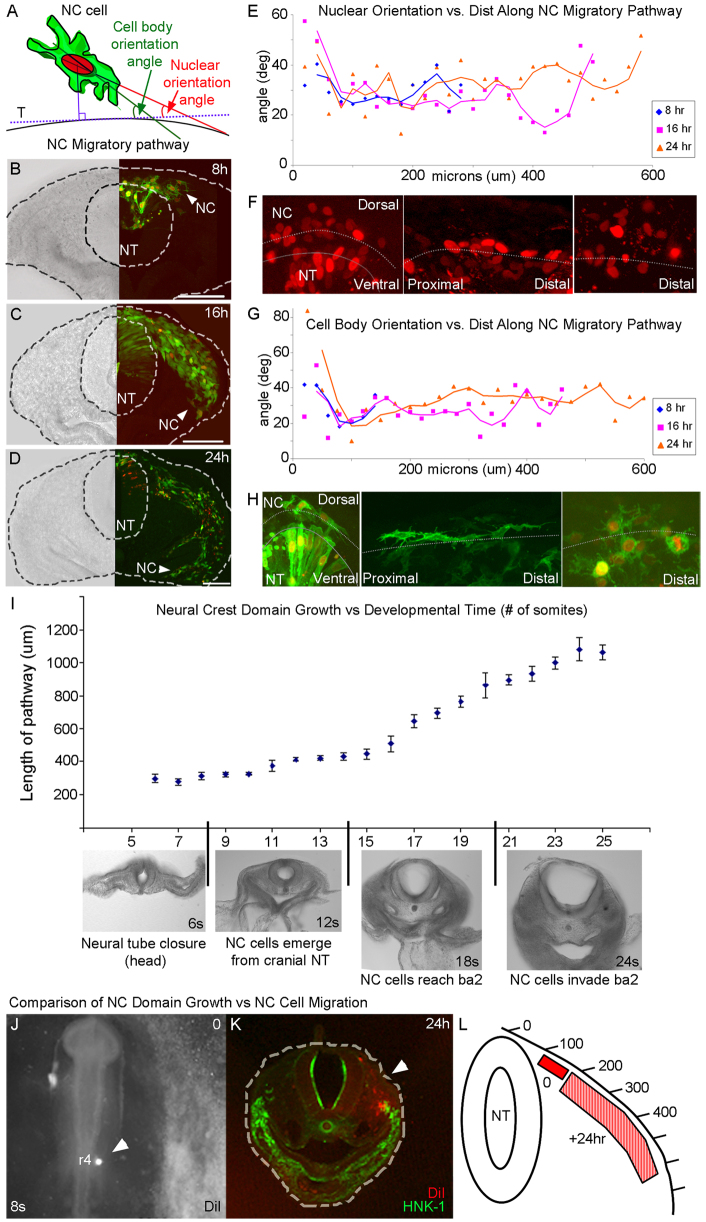
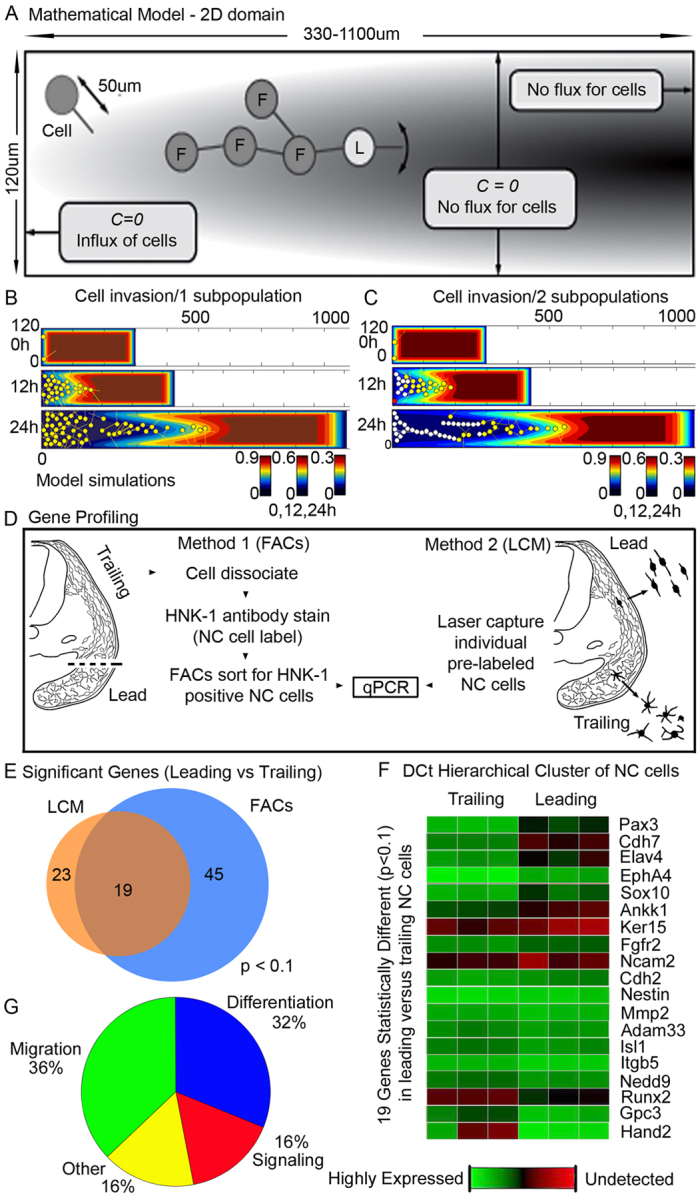
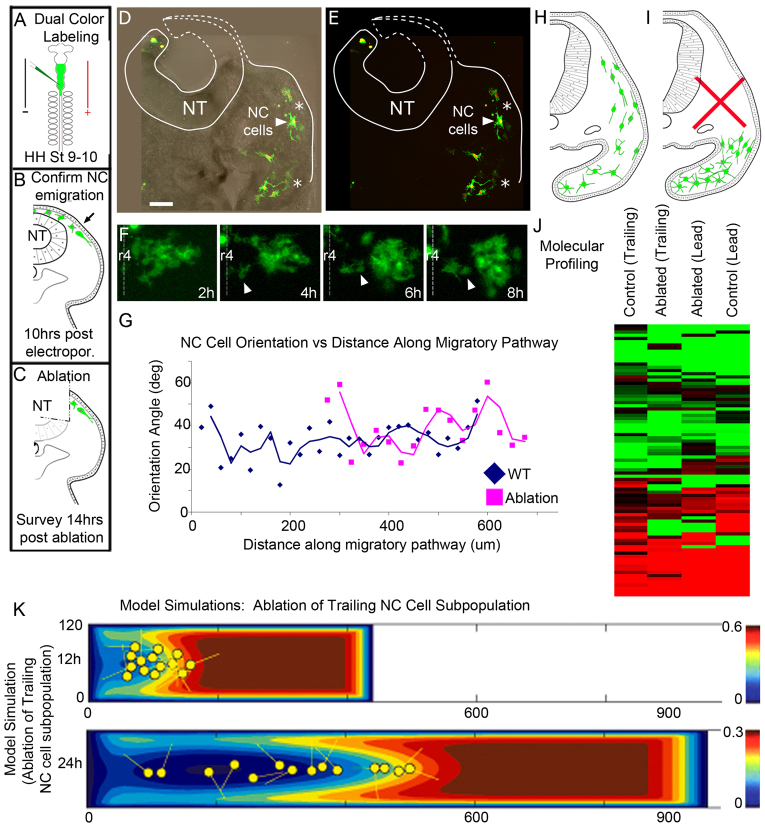
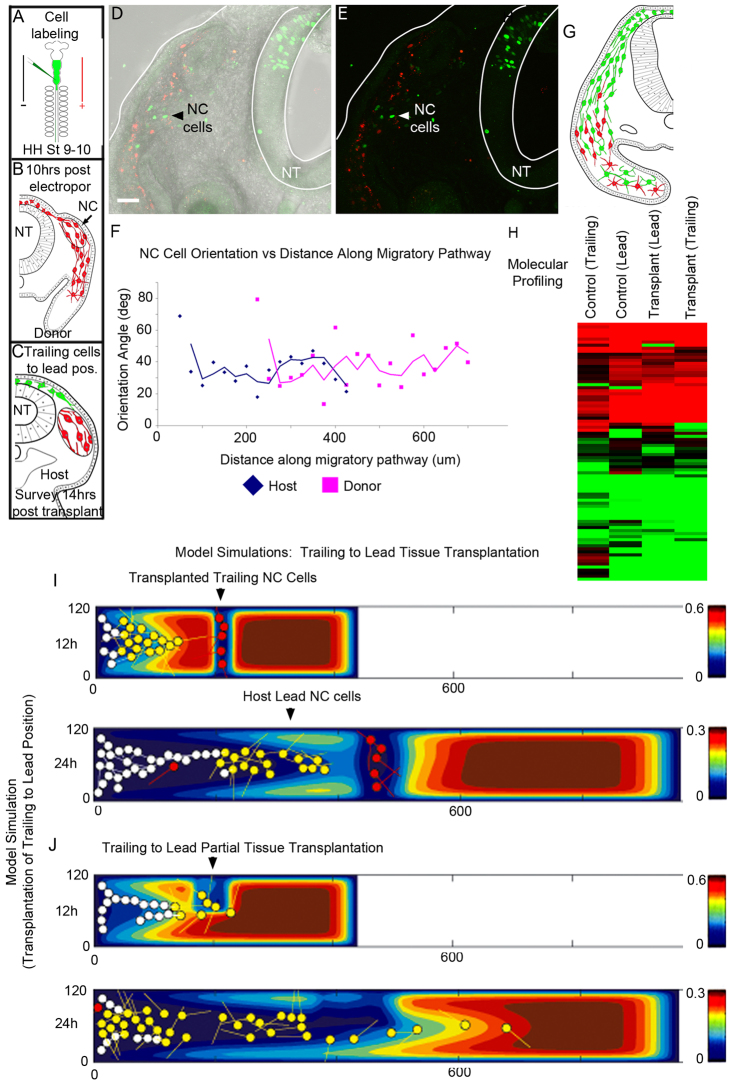
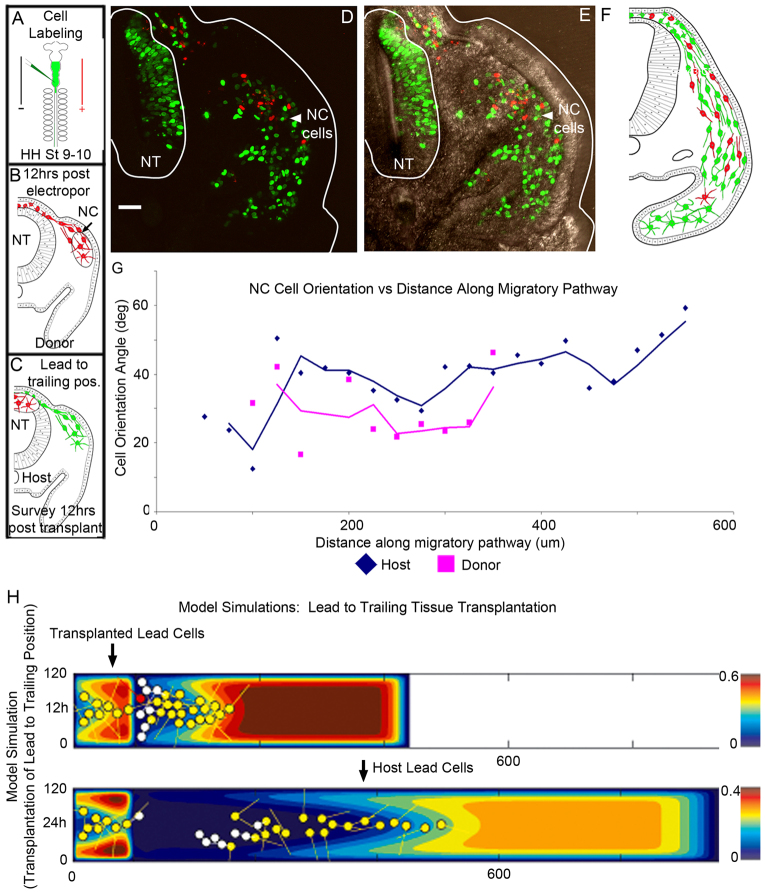
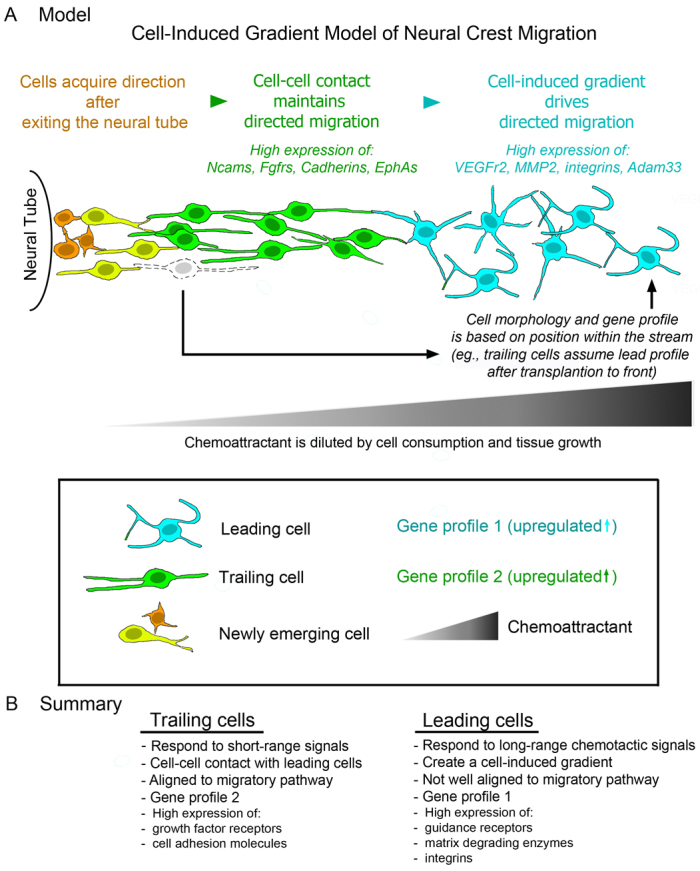
Long-distance cell migration is an important feature of embryonic development, adult morphogenesis and cancer, yet the mechanisms that drive subpopulations of cells to distinct targets are poorly understood. Here, we use the embryonic neural crest (NC) in tandem with theoretical studies to evaluate model mechanisms of long-distance cell migration. We find that a simple chemotaxis model is insufficient to explain our experimental data. Instead, model simulations predict that NC cell migration requires leading cells to respond to long-range guidance signals and trailing cells to short-range cues in order to maintain a directed, multicellular stream. Experiments confirm differences in leading versus trailing NC cell subpopulations, manifested in unique cell orientation and gene expression patterns that respond to non-linear tissue growth of the migratory domain. Ablation experiments that delete the trailing NC cell subpopulation reveal that leading NC cells distribute all along the migratory pathway and develop a leading/trailing cellular orientation and gene expression profile that is predicted by model simulations. Transplantation experiments and model predictions that move trailing NC cells to the migratory front, or vice versa, reveal that cells adopt a gene expression profile and cell behaviors corresponding to the new position within the migratory stream. These results offer a mechanistic model in which leading cells create and respond to a cell-induced chemotactic gradient and transmit guidance information to trailing cells that use short-range signals to move in a directional manner.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
