

Molecular characterization of proprotein convertase subtilisin/kexin type 9-mediated degradation of the LDLR
- PMID: 22764087
- PMCID: PMC3413232
- DOI: 10.1194/jlr.M028563
Molecular characterization of proprotein convertase subtilisin/kexin type 9-mediated degradation of the LDLR
Abstract
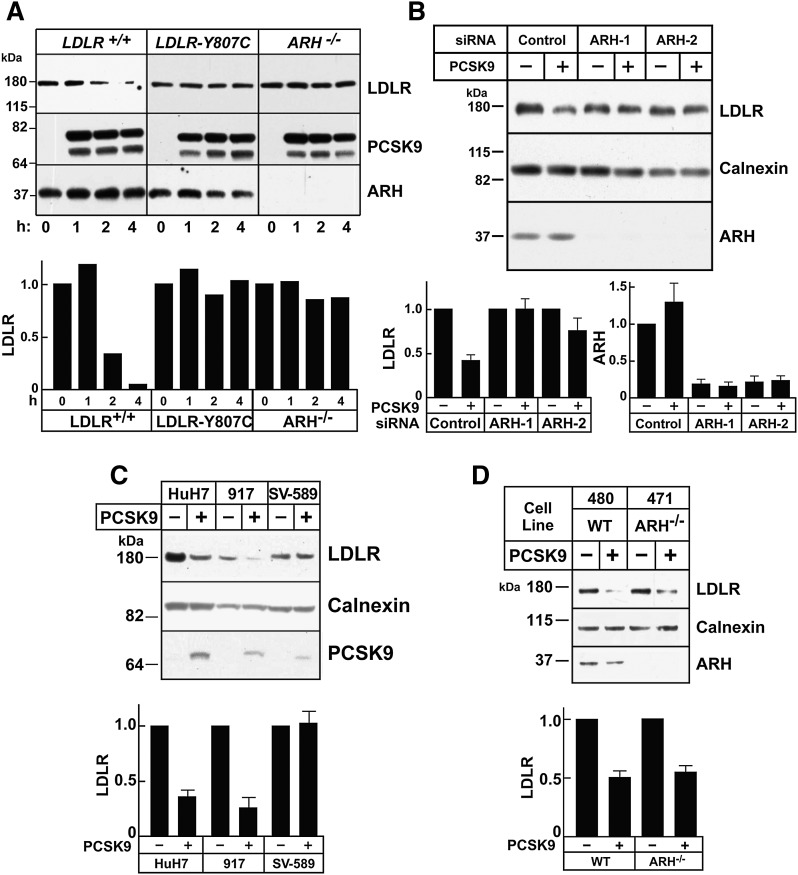
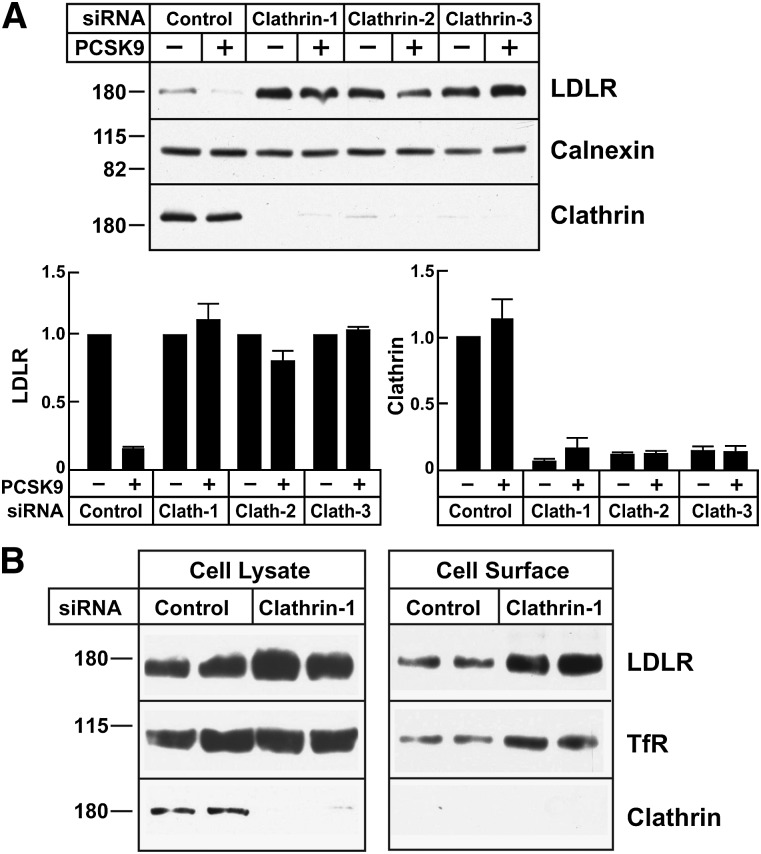
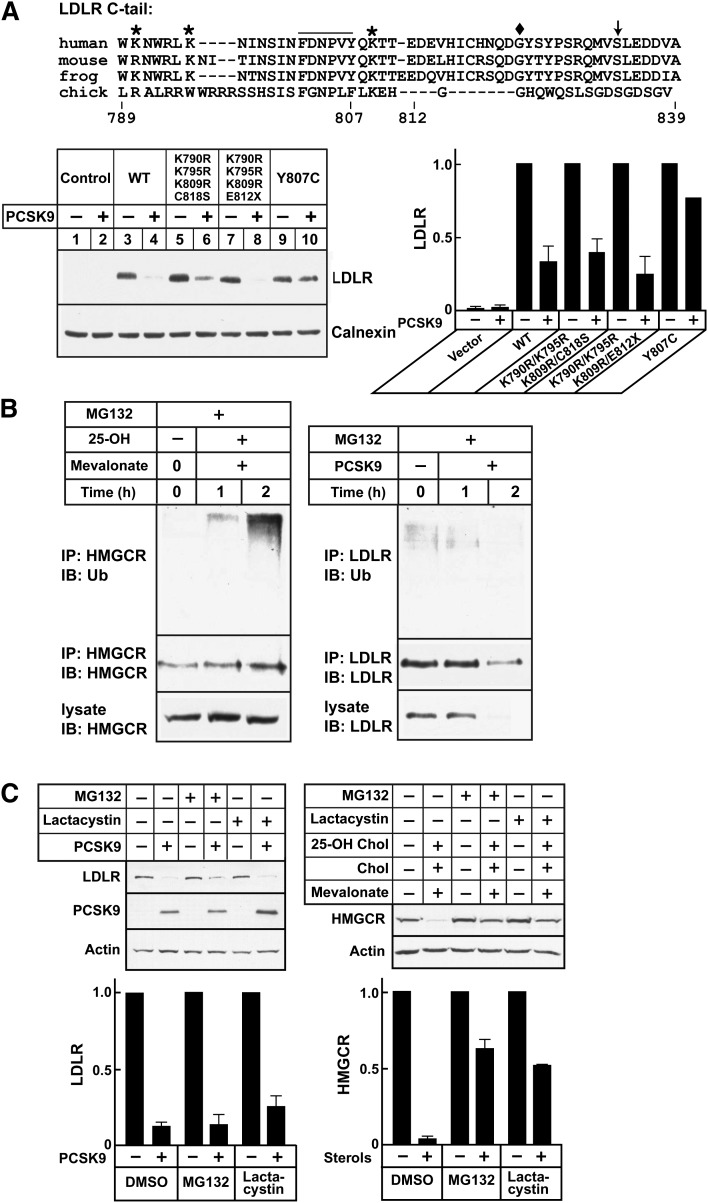
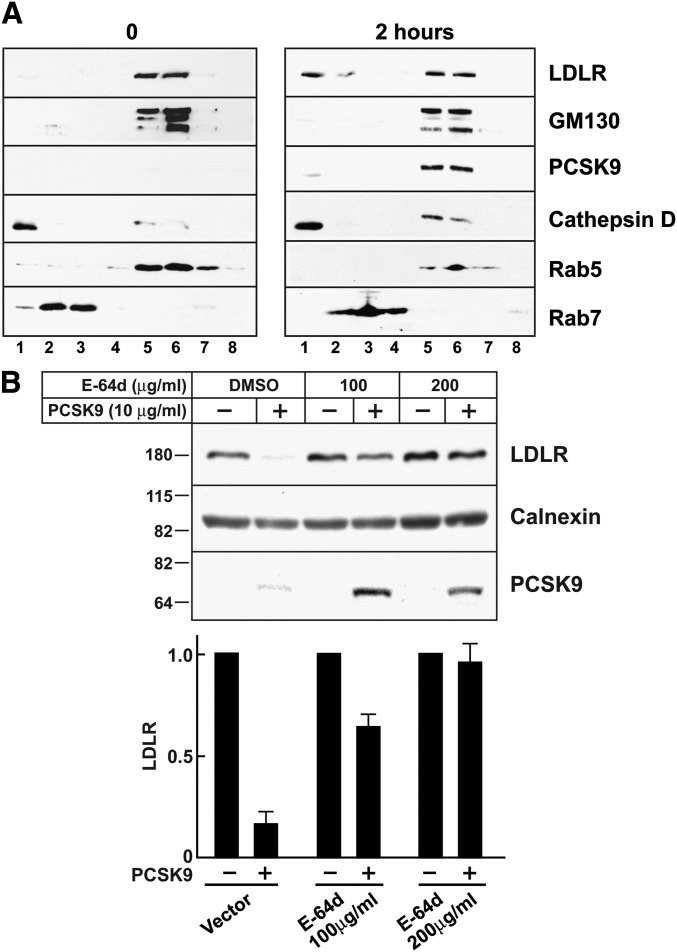
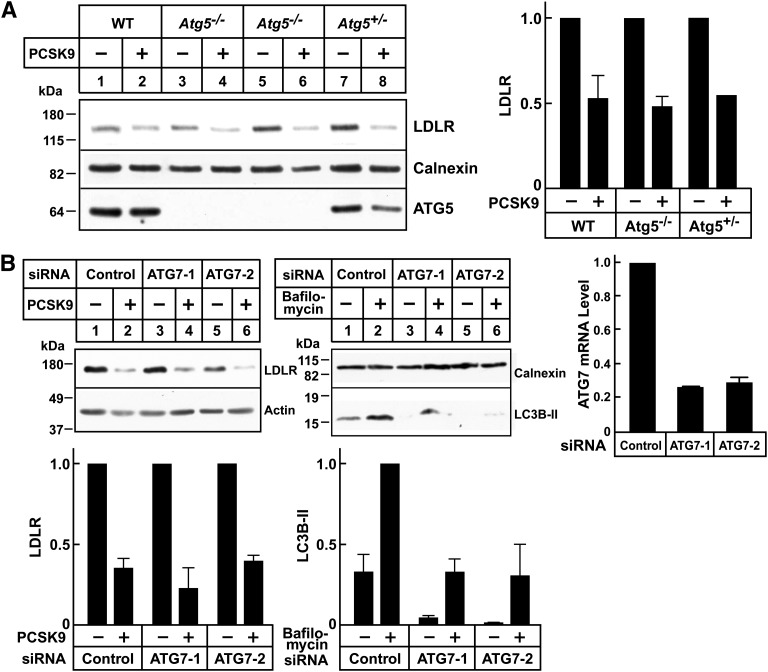
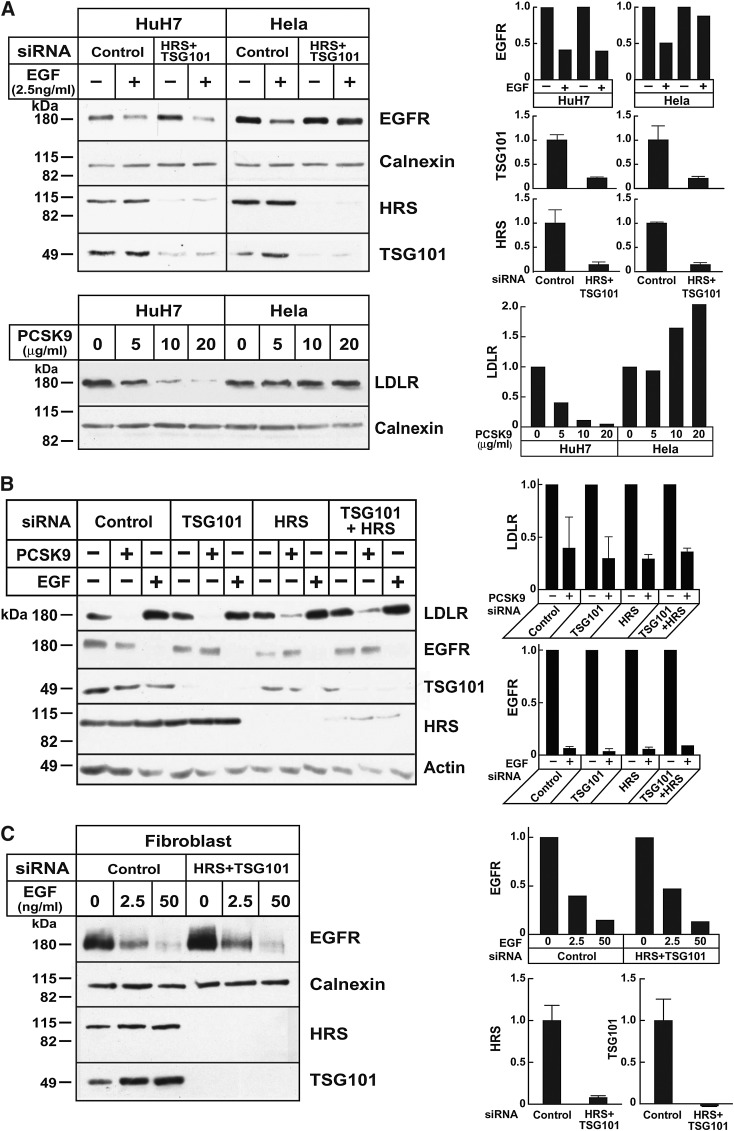
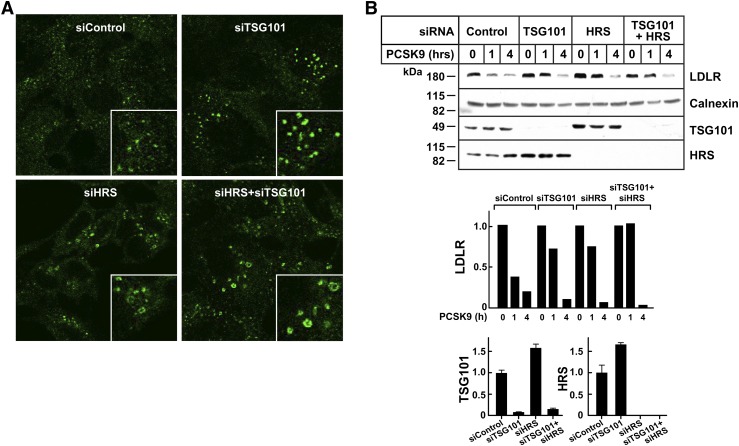
Proprotein convertase subtilisin/kexin type 9 (PCSK9) is a secreted protein that promotes degradation of cell surface LDL receptors (LDLRs) in selected cell types. Here we used genetic and pharmacological inhibitors to define the pathways involved in PCSK9-mediated LDLR degradation. Inactivating mutations in autosomal recessive hypercholesterolemia (ARH), an endocytic adaptor, blocked PCSK9-mediated LDLR degradation in lymphocytes but not in fibroblasts. Thus, ARH is not specifically required for PCSK9-mediated LDLR degradation. Knockdown of clathrin heavy chain with siRNAs prevented LDLR degradation. In contrast, prevention of ubiquitination of the LDLR cytoplasmic tail, inhibition of proteasomal activity, or disruption of proteins required for lysosomal targeting via macroautophagy (autophagy related 5 and 7) or the endosomal sorting complex required for trafficking (ESCRT) pathway (hepatocyte growth factor-regulated Tyr-kinase substrate and tumor suppressor gene 101) failed to block PCSK9-mediated LDLR degradation. These findings are consistent with a model in which the LDLR-PCSK9 complex is internalized via clathrin-mediated endocytosis and then routed to lysosomes via a mechanism that does not require ubiquitination and is distinct from the autophagy and proteosomal degradation pathways. Finally, the PCSK9-LDLR complex appears not to be transported by the canonical ESCRT pathway.
Figures
References
-
- Brown M. S., Goldstein J. L. 1986. A receptor-mediated pathway for cholesterol homeostasis. Science. 232: 34–47. - PubMed
-
- Park S. W., Moon Y. A., Horton J. D. 2004. Post-transcriptional regulation of low density lipoprotein receptor protein by proprotein convertase subtilisin/kexin type 9a in mouse liver. J. Biol. Chem. 279: 50630–50638. - PubMed
-
- Benjannet S., Rhainds D., Essalmani R., Mayne J., Wickham L., Jin W., Asselin M. C., Hamelin J., Varret M., Allard D., et al. 2004. NARC-1/PCSK9 and its natural mutants: zymogen cleavage and effects on the LDLR and LDL-cholesterol. J. Biol. Chem. 279: 48865–48875. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
