

A Lys49-PLA2 myotoxin of Bothrops asper triggers a rapid death of macrophages that involves autocrine purinergic receptor signaling
- PMID: 22764102
- PMCID: PMC3406575
- DOI: 10.1038/cddis.2012.68
A Lys49-PLA2 myotoxin of Bothrops asper triggers a rapid death of macrophages that involves autocrine purinergic receptor signaling
Abstract
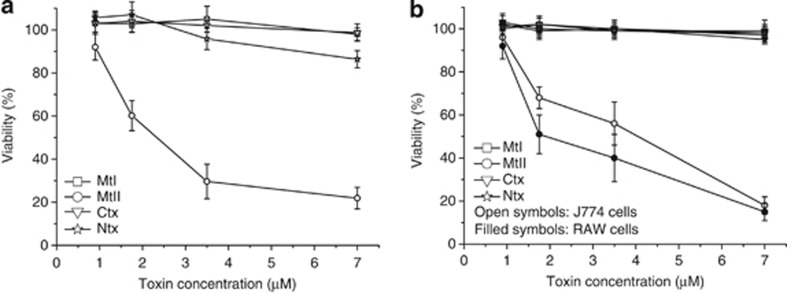
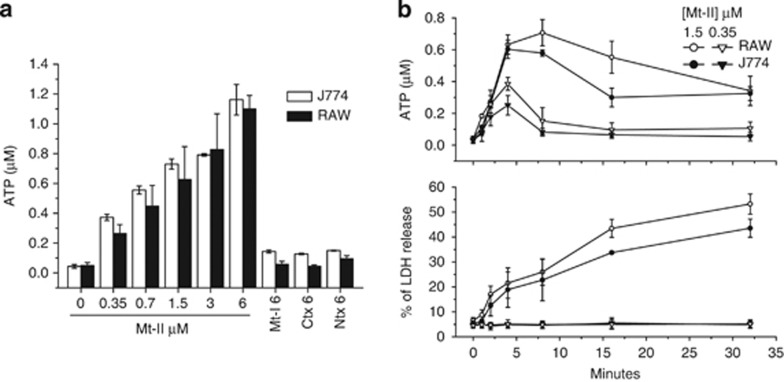
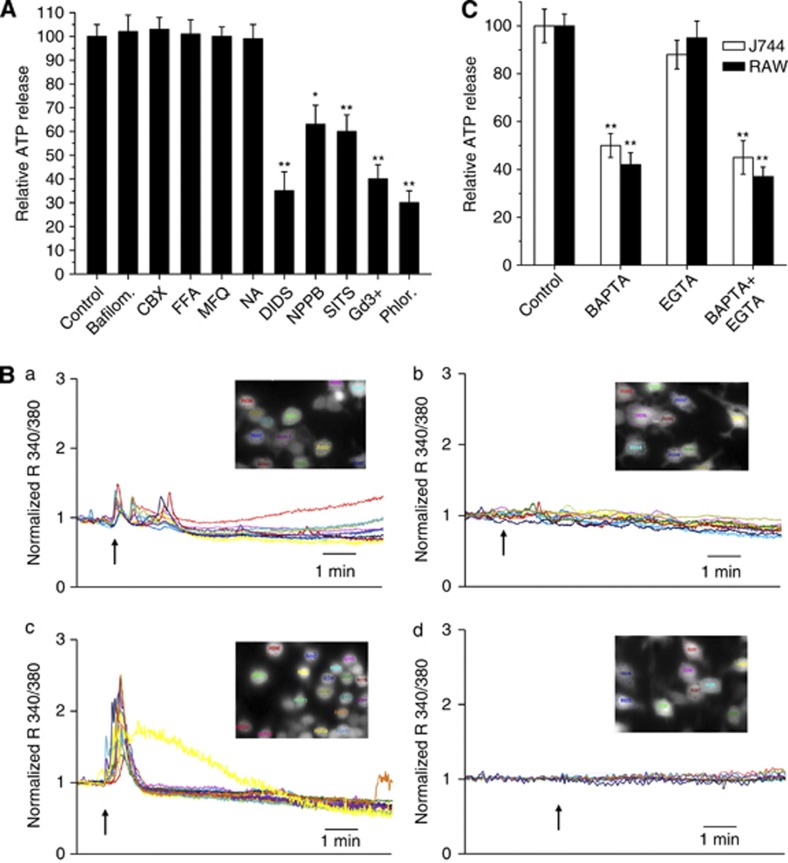
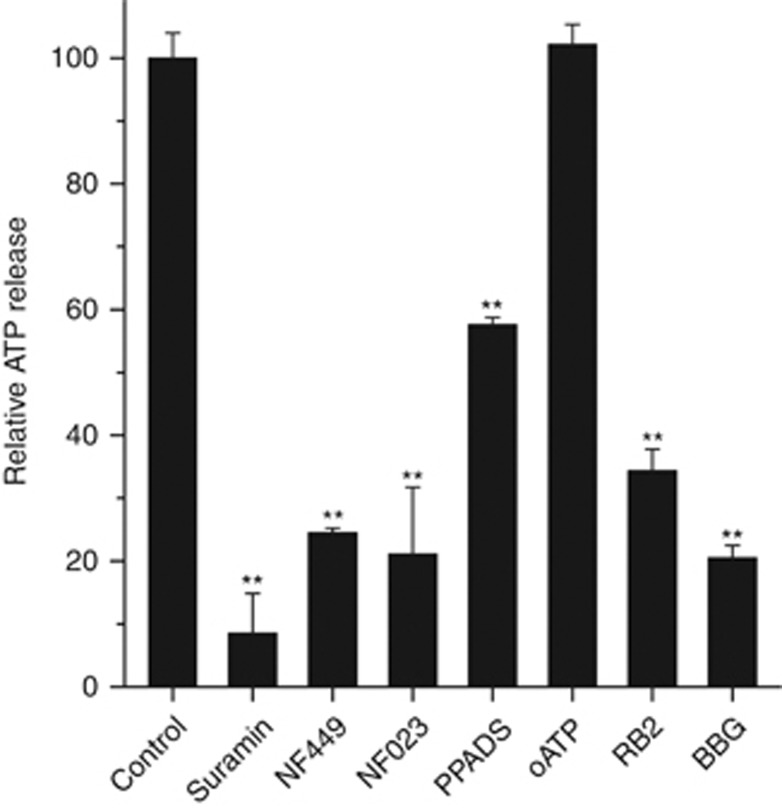
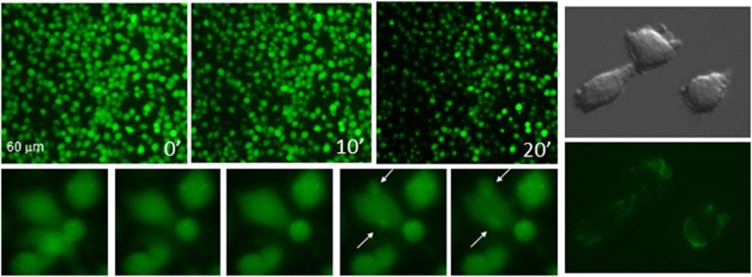
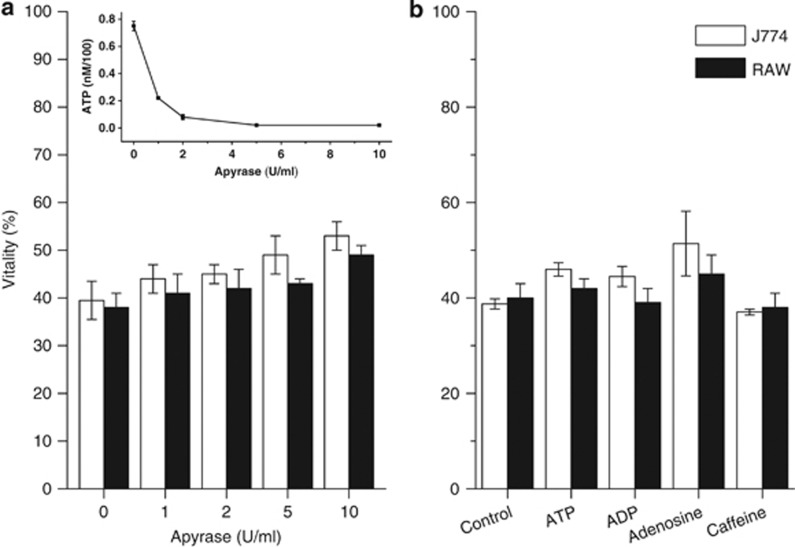
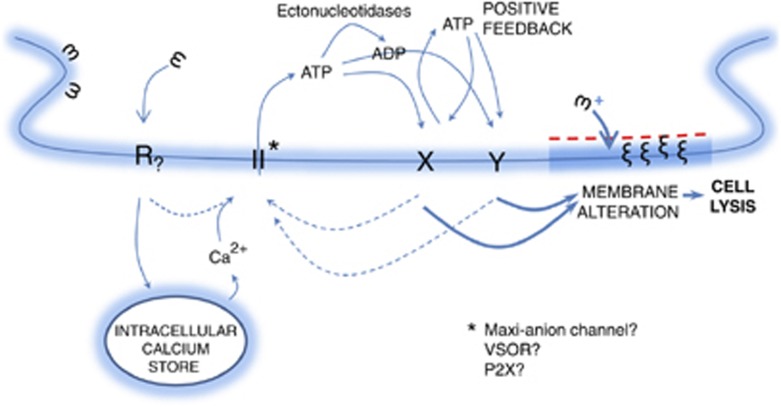
Lys49-PLA(2) myotoxins, an important component of various viperid snake venoms, are a class of PLA(2)-homolog proteins deprived of catalytic activity. Similar to enzymatically active PLA(2) (Asp49) and to other classes of myotoxins, they cause severe myonecrosis. Moreover, these toxins are used as tools to study skeletal muscle repair and regeneration, a process that can be very limited after snakebites. In this work, the cytotoxic effect of different myotoxins, Bothrops asper Lys49 and Asp49-PLA(2), Notechis scutatus notexin and Naja mossambica cardiotoxin, was evaluated on macrophages, cells that have a key role in muscle regeneration. Only the Lys49-myotoxin was found to trigger a rapid asynchronous death of mouse peritoneal macrophages and macrophagic cell lines through a process that involves ATP release, ATP-induced ATP release and that is inhibited by various purinergic receptor antagonists. ATP leakage is induced also at sublytical doses of the Lys49-myotoxin, it involves Ca(2+) release from intracellular stores, and is reduced by inhibitors of VSOR and the maxi-anion channel. The toxin-induced cell death is different from that caused by high concentration of ATP and appears to be linked to localized purinergic signaling. Based on present findings, a mechanism of cell death is proposed that can be extended to other cytolytic proteins and peptides.
Figures
References
-
- Gutiérrez JM, Rucavado A, Chaves F, Díaz C, Escalante T. Experimental pathology of local tissue damage induced by Bothrops asper snake venom. Toxicon. 2009;54:958–975. - PubMed
-
- Gutiérrez JM, Escalante T, Rucavado A. Experimental pathophysiology of systemic alterations induced by Bothrops asper snake venom. Toxicon. 2009;54:976–987. - PubMed
-
- Lomonte B, Angulo Y, Calderón L. An overview of lysine-49 phospholipase A2 myotoxins from crotalid snake venoms and their structural determinants of myotoxic action. Toxicon. 2003;42:885–901. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
