

Otx2 binding to perineuronal nets persistently regulates plasticity in the mature visual cortex
- PMID: 22764251
- PMCID: PMC3419577
- DOI: 10.1523/JNEUROSCI.0394-12.2012
Otx2 binding to perineuronal nets persistently regulates plasticity in the mature visual cortex
Abstract
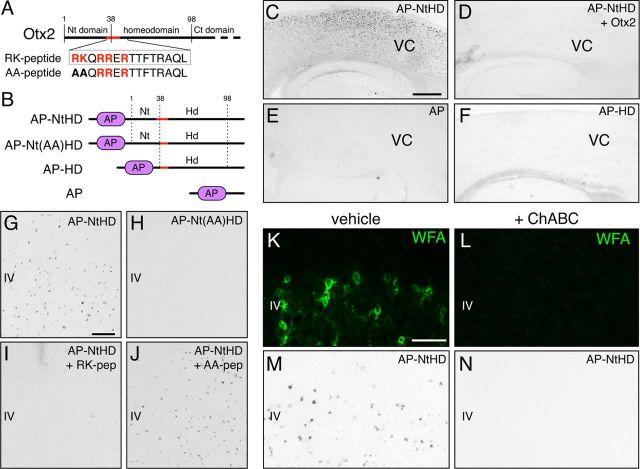
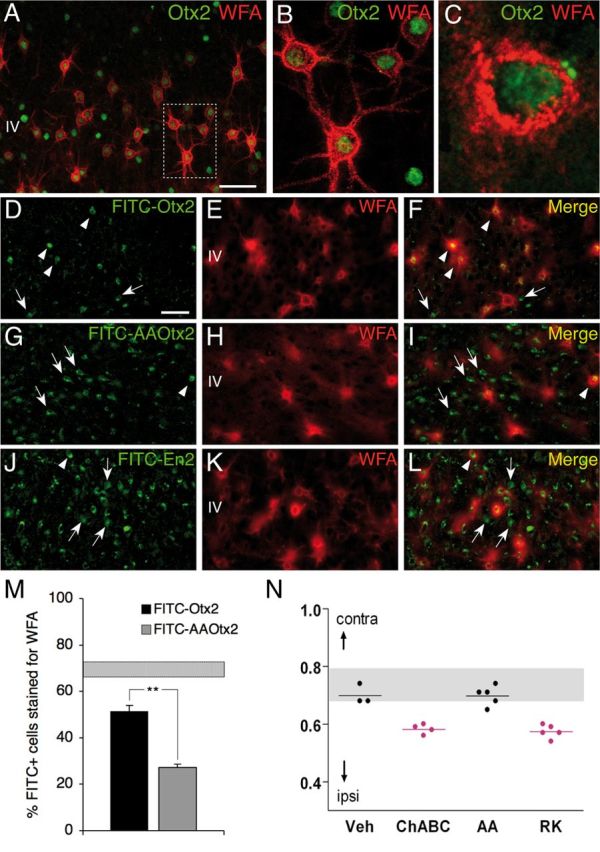
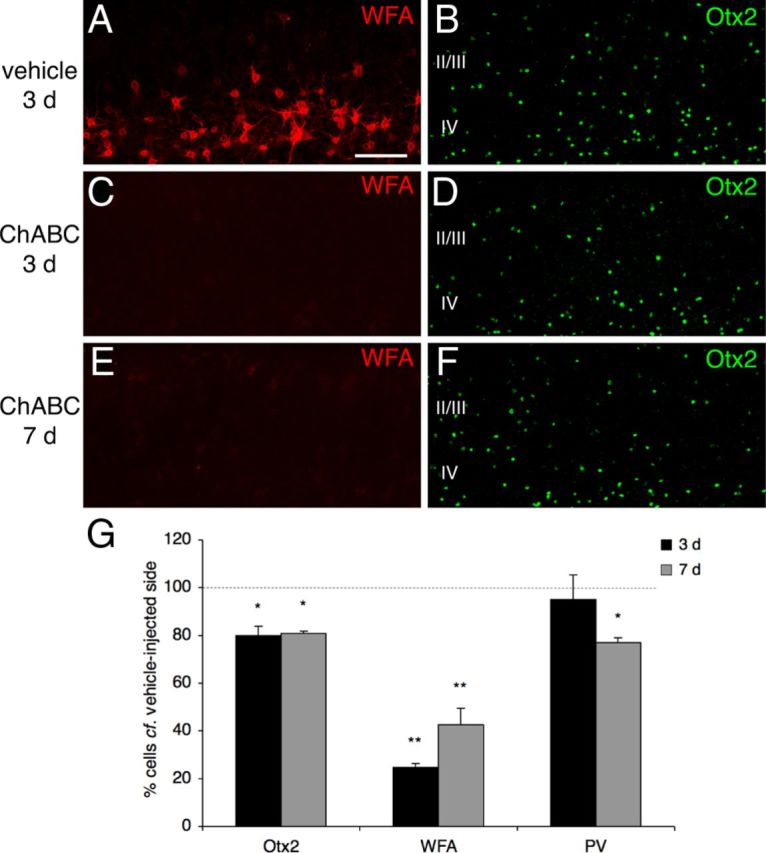
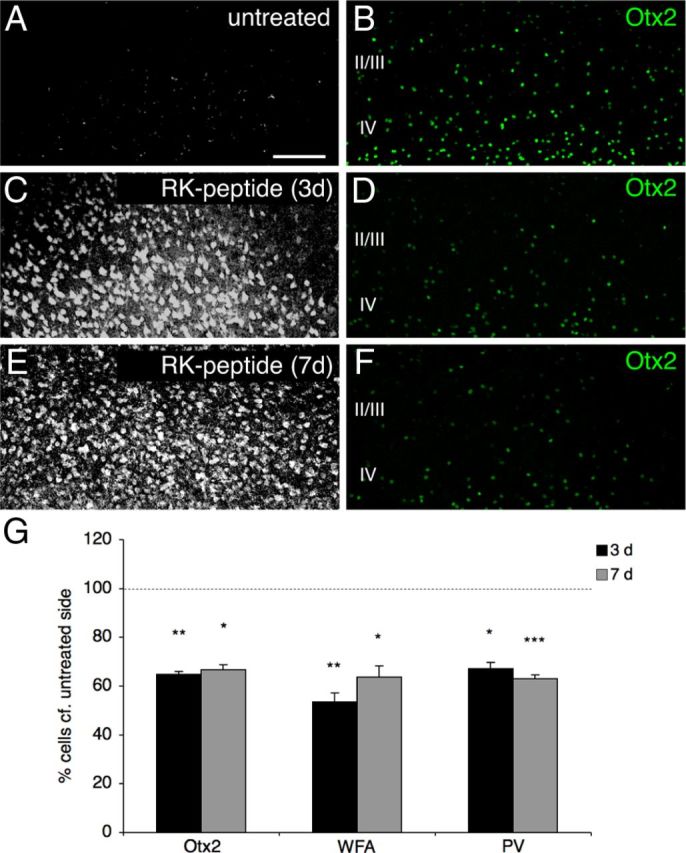
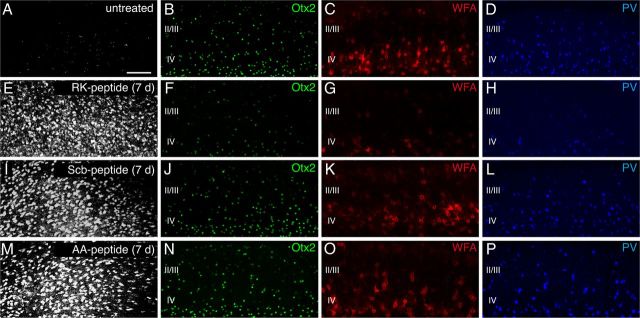
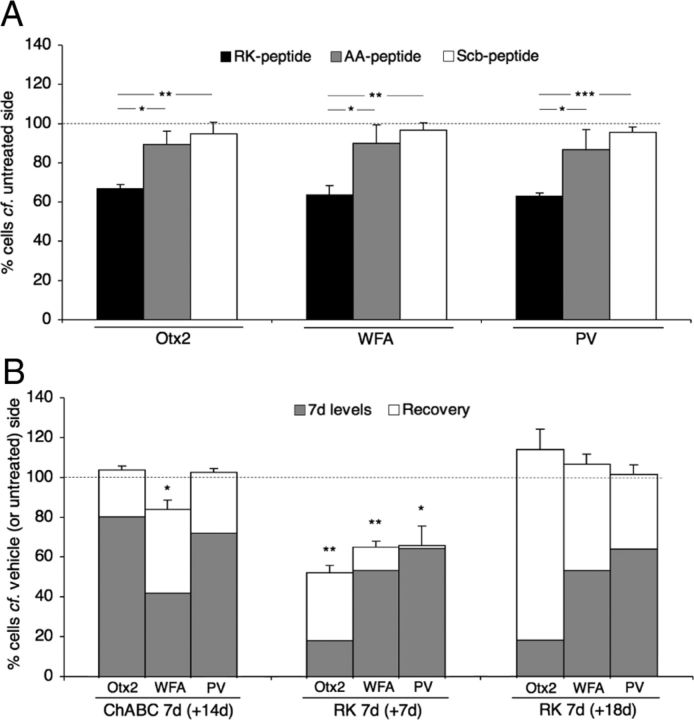
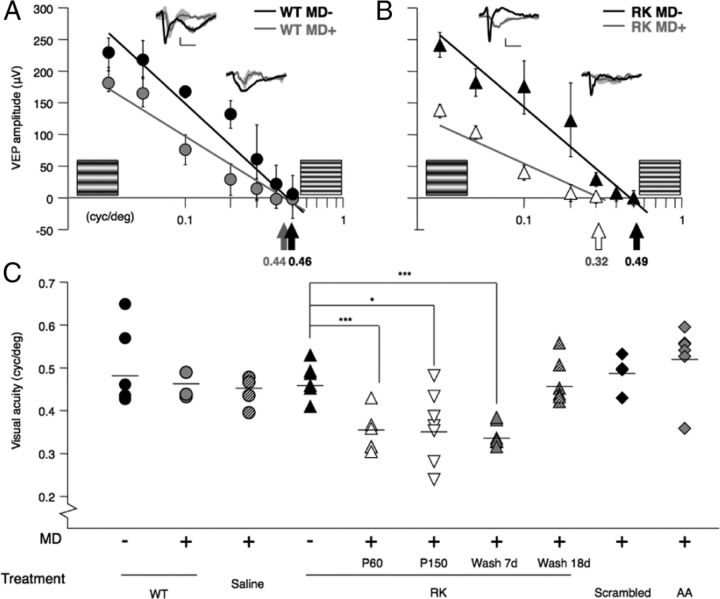
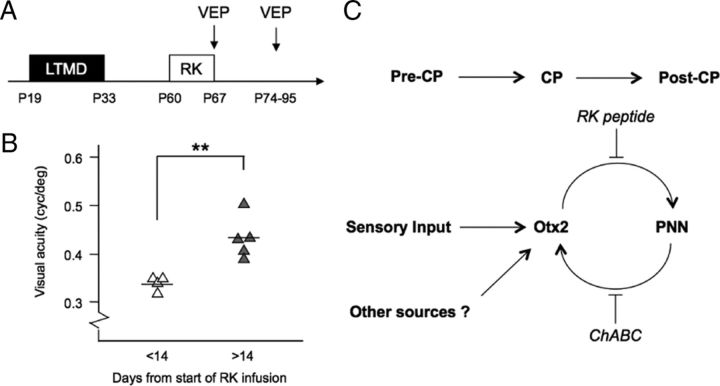
Specific transfer of (orthodenticle homeobox 2) Otx2 homeoprotein into GABAergic interneurons expressing parvalbumin (PV) is necessary and sufficient to open, then close, a critical period (CP) of plasticity in the developing mouse visual cortex. The accumulation of endogenous Otx2 in PV cells suggests the presence of specific Otx2 binding sites. Here, we find that perineuronal nets (PNNs) on the surfaces of PV cells permit the specific, constitutive capture of Otx2. We identify a 15 aa domain containing an arginine-lysine doublet (RK peptide) within Otx2, bearing prototypic traits of a glycosaminoglycan (GAG) binding sequence that mediates Otx2 binding to PNNs, and specifically to chondroitin sulfate D and E, with high affinity. Accordingly, PNN hydrolysis by chondroitinase ABC reduces the amount of endogenous Otx2 in PV cells. Direct infusion of RK peptide similarly disrupts endogenous Otx2 localization to PV cells, reduces PV and PNN expression, and reopens plasticity in adult mice. The closure of one eye during this transient window reduces cortical acuity and is specific to the RK motif, as an Alanine-Alanine variant or a scrambled peptide fails to reactivate plasticity. Conversely, this transient reopening of plasticity in the adult restores binocular vision in amblyopic mice. Thus, one function of PNNs is to facilitate the persistent internalization of Otx2 by PV cells to maintain CP closure. The pharmacological use of the Otx2 GAG binding domain offers a novel, potent therapeutic tool with which to restore cortical plasticity in the mature brain.
Figures
References
-
- Berardi N, Pizzorusso T, Maffei L. Extracellular matrix and visual cortical plasticity: freeing the synapse. Neuron. 2004;44:905–908. - PubMed
-
- Brunet I, Di Nardo AA, Sonnier L, Beurdeley M, Prochiantz A. The topological role of homeoproteins in the developing central nervous system. Trends Neurosci. 2007;30:260–267. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous