

Cdk1-dependent control of membrane-trafficking dynamics
- PMID: 22767578
- PMCID: PMC3431941
- DOI: 10.1091/mbc.E11-10-0834
Cdk1-dependent control of membrane-trafficking dynamics
Abstract
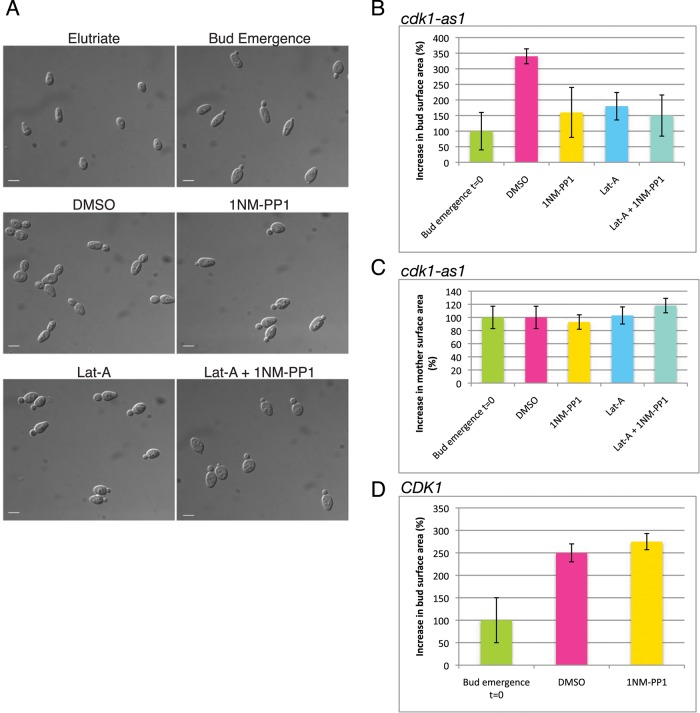
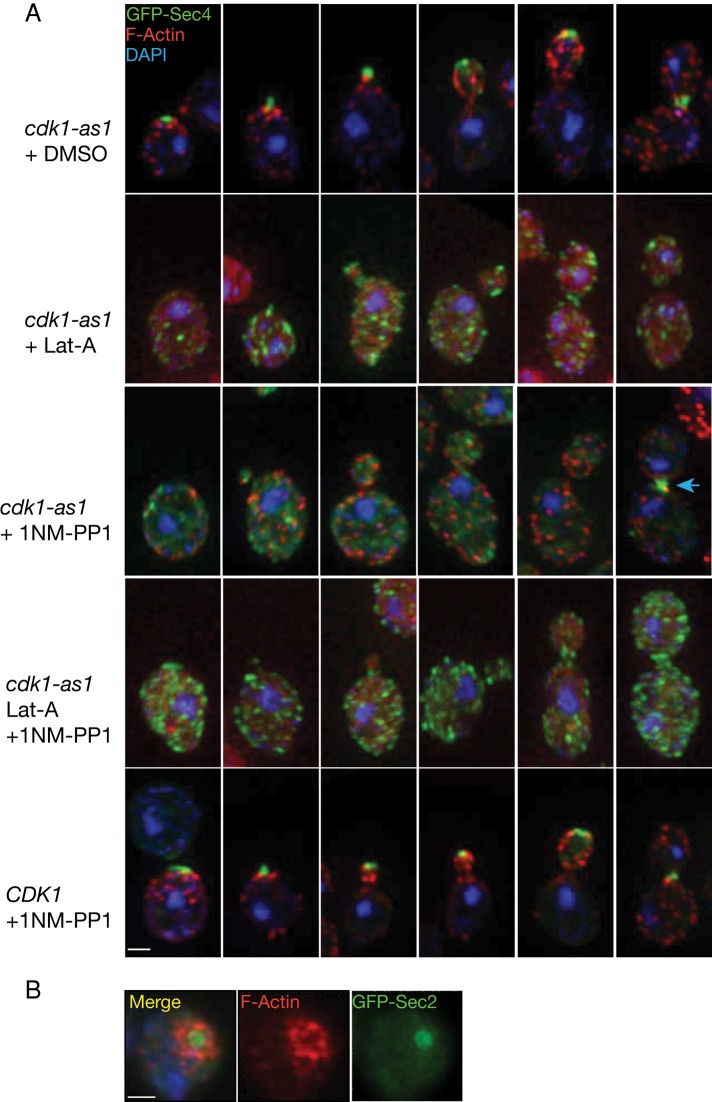
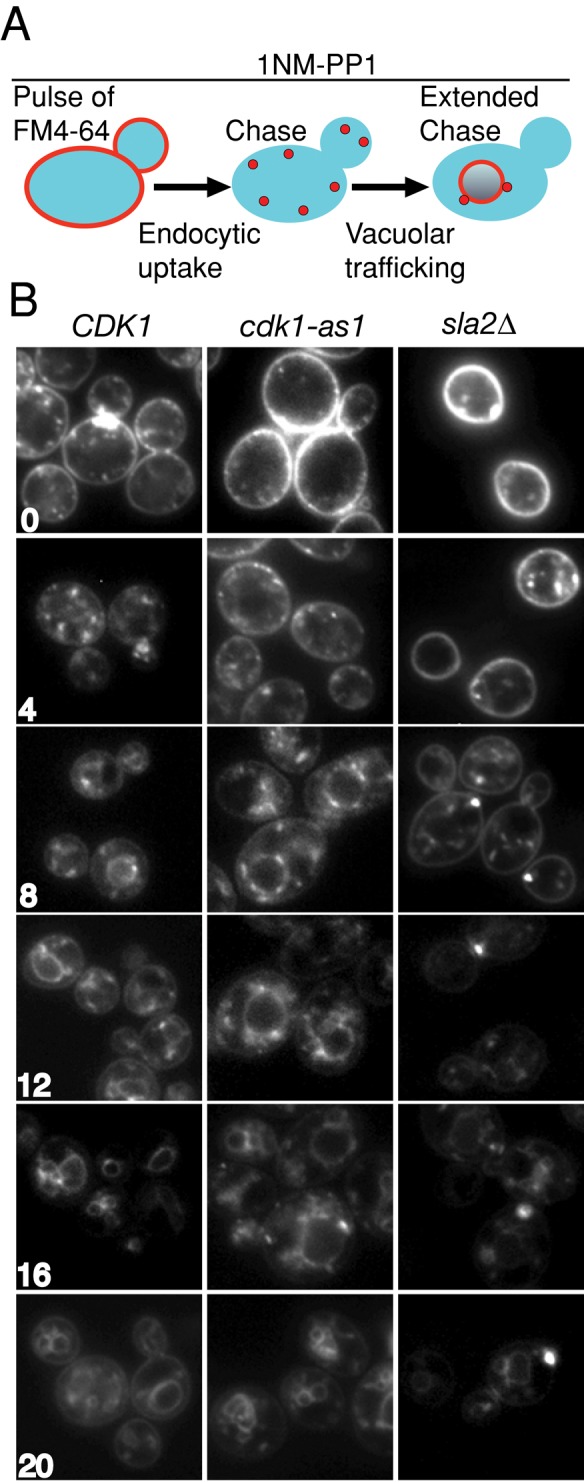
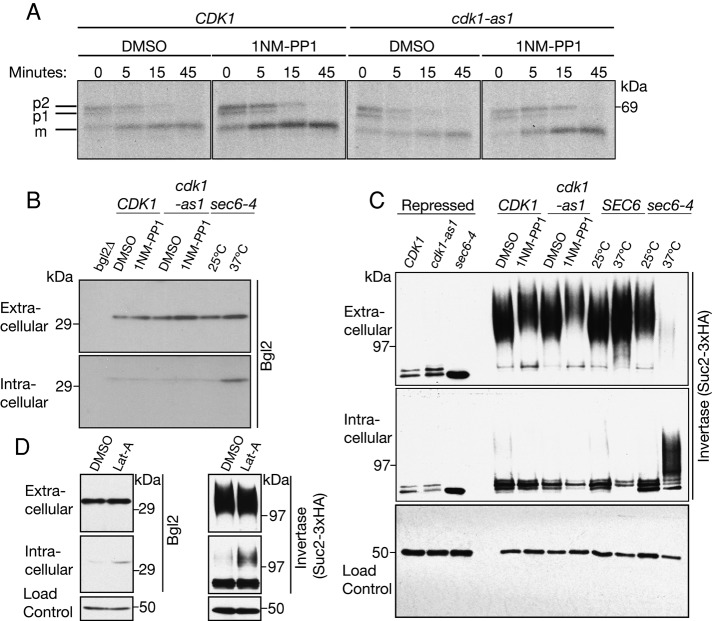
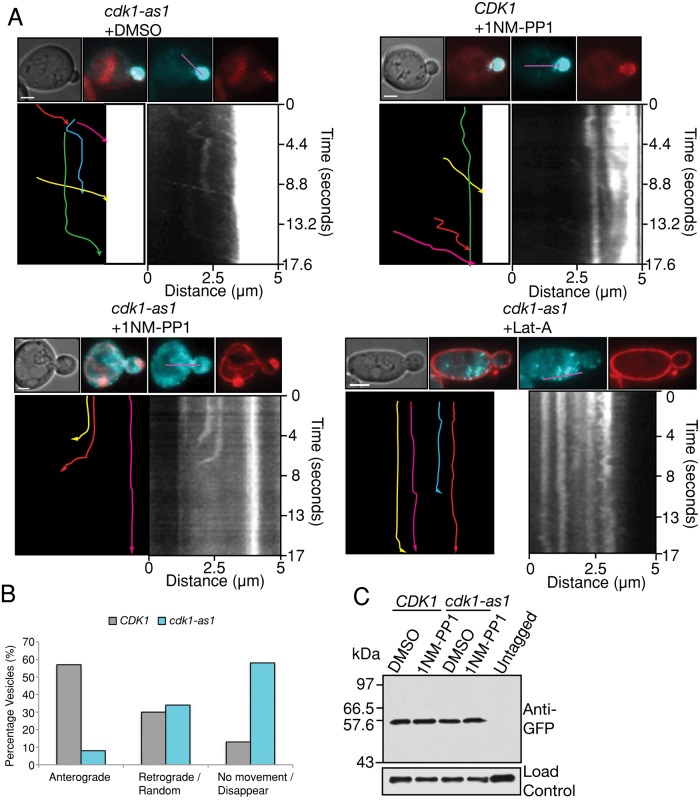
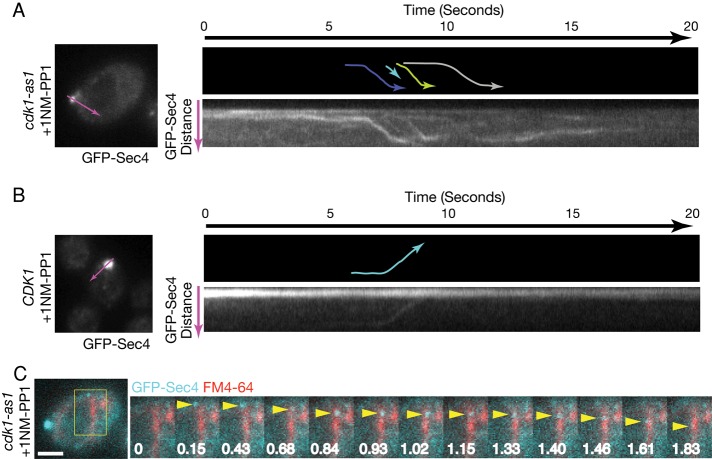
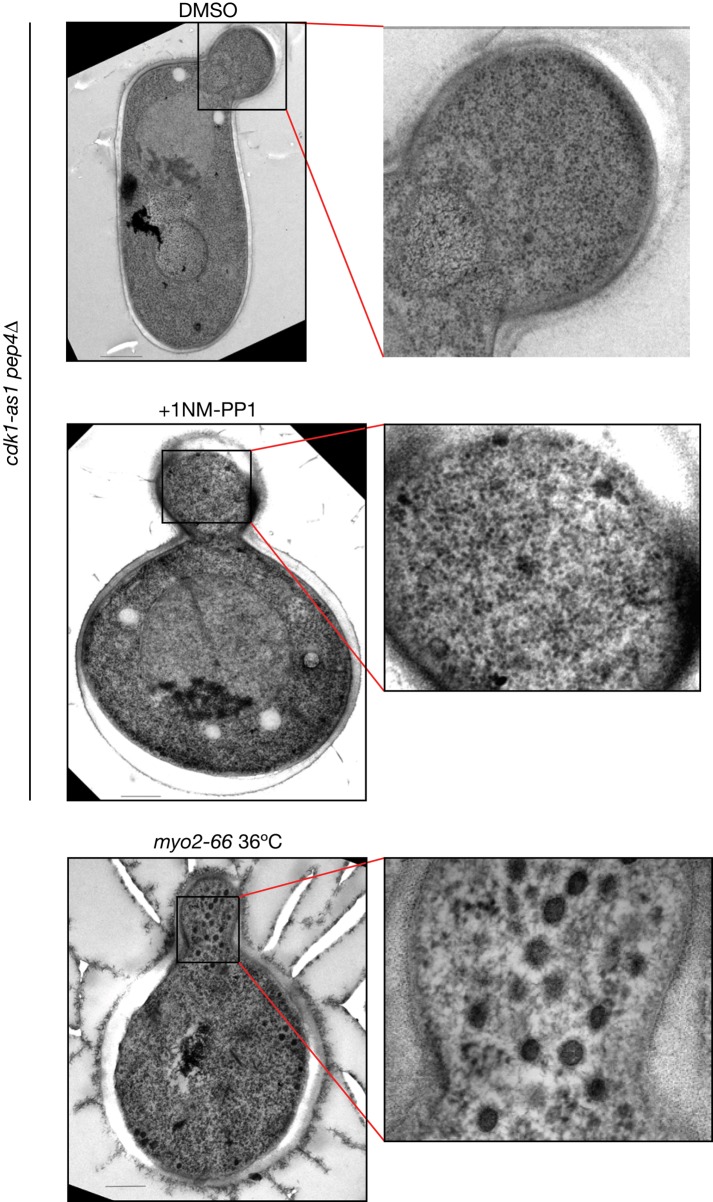
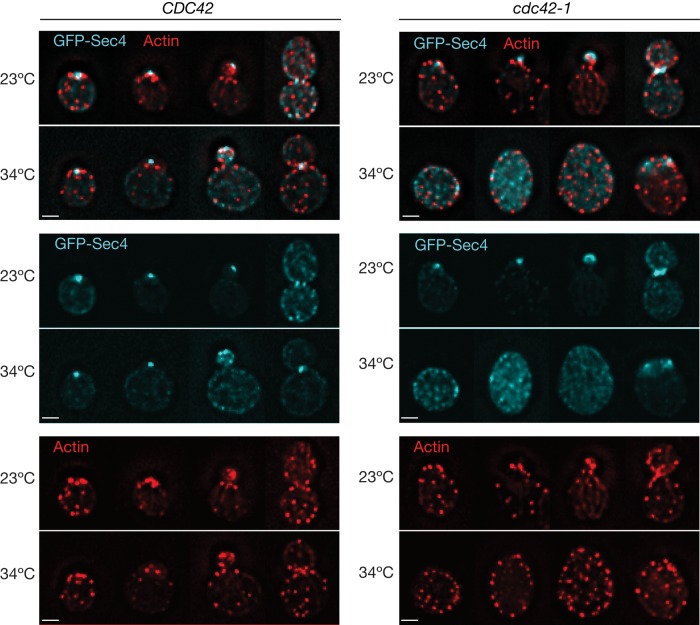
Cyclin-dependent kinase 1 (Cdk1) is required for initiation and maintenance of polarized cell growth in budding yeast. Cdk1 activates Rho-family GTPases, which polarize the actin cytoskeleton for delivery of membrane to growth sites via the secretory pathway. Here we investigate whether Cdk1 plays additional roles in the initiation and maintenance of polarized cell growth. We find that inhibition of Cdk1 causes a cell surface growth defect that is as severe as that caused by actin depolymerization. However, unlike actin depolymerization, Cdk1 inhibition does not result in a massive accumulation of intracellular secretory vesicles or their cargoes. Analysis of post-Golgi vesicle dynamics after Cdk1 inhibition demonstrates that exocytic vesicles are rapidly mistargeted away from the growing bud, possibly to the endomembrane/vacuolar system. Inhibition of Cdk1 also causes defects in the organization of endocytic and exocytic zones at the site of growth. Cdk1 thus modulates membrane-trafficking dynamics, which is likely to play an important role in coordinating cell surface growth with cell cycle progression.
Figures
Similar articles
-
Cdk1 coordinates cell-surface growth with the cell cycle.Nat Cell Biol. 2007 May;9(5):506-15. doi: 10.1038/ncb1568. Epub 2007 Apr 8. Nat Cell Biol. 2007. PMID: 17417630
-
Yeast Cdc42 functions at a late step in exocytosis, specifically during polarized growth of the emerging bud.J Cell Biol. 2001 Nov 12;155(4):581-92. doi: 10.1083/jcb.200106065. Epub 2001 Nov 12. J Cell Biol. 2001. PMID: 11706050 Free PMC article.
-
Secretory pathway-dependent localization of the Saccharomyces cerevisiae Rho GTPase-activating protein Rgd1p at growth sites.Eukaryot Cell. 2012 May;11(5):590-600. doi: 10.1128/EC.00042-12. Epub 2012 Mar 23. Eukaryot Cell. 2012. PMID: 22447923 Free PMC article.
-
Spatiotemporal Control of Intracellular Membrane Trafficking by Rho GTPases.Cells. 2019 Nov 21;8(12):1478. doi: 10.3390/cells8121478. Cells. 2019. PMID: 31766364 Free PMC article. Review.
-
Rho GTPases: regulation of cell polarity and growth in yeasts.Biochem J. 2010 Feb 24;426(3):243-53. doi: 10.1042/BJ20091823. Biochem J. 2010. PMID: 20175747 Review.
Cited by
-
The Trypanosome Exocyst: A Conserved Structure Revealing a New Role in Endocytosis.PLoS Pathog. 2017 Jan 23;13(1):e1006063. doi: 10.1371/journal.ppat.1006063. eCollection 2017 Jan. PLoS Pathog. 2017. PMID: 28114397 Free PMC article.
-
Plasma membrane growth during the cell cycle: unsolved mysteries and recent progress.Curr Opin Cell Biol. 2012 Dec;24(6):845-51. doi: 10.1016/j.ceb.2012.10.008. Epub 2012 Nov 7. Curr Opin Cell Biol. 2012. PMID: 23141634 Free PMC article. Review.
-
Roles of the fission yeast UNC-13/Munc13 protein Ync13 in late stages of cytokinesis.Mol Biol Cell. 2018 Sep 15;29(19):2259-2279. doi: 10.1091/mbc.E18-04-0225. Epub 2018 Jul 25. Mol Biol Cell. 2018. PMID: 30044717 Free PMC article.
-
Cell Size and Growth Rate Are Modulated by TORC2-Dependent Signals.Curr Biol. 2018 Jan 22;28(2):196-210.e4. doi: 10.1016/j.cub.2017.11.069. Epub 2017 Dec 28. Curr Biol. 2018. PMID: 29290562 Free PMC article.
-
Mitotic phosphorylation of Exo84 disrupts exocyst assembly and arrests cell growth.J Cell Biol. 2013 Jul 8;202(1):97-111. doi: 10.1083/jcb.201211093. J Cell Biol. 2013. PMID: 23836930 Free PMC article.
References
-
- Bishop AC, Buzko O, Shokat KM. Magic bullets for protein kinases. Trends Cell Biol. 2001;11:167–172. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
